
树突状细胞(DC)是一组不同类型的造血细胞,在先天免疫系统和后天免疫系统之间起着通道的作用。它们起源于淋巴-骨髓造血作用,来源于骨髓。它们是先天免疫细胞,因为它们可以识别病原体,但它们也可以在主要组织相容性复合体(MHC)蛋白的作用下制备和呈递抗原,以激发初始T细胞对威胁作出应答。已确认至少三种树突状细胞:浆细胞样DC(pDC)和髓样DC /传统DC(cDC)。1它们在肿瘤微环境中起关键作用。2有很多研究利用DC开发癌症、慢性感染和自身免疫性疾病的免疫疗法以及诱导移植耐受。BD继续扩大其仪器和试剂组合,以便用多色流式细胞仪对DC及其不同亚群进行富集、分选和分析。
树突状细胞生物学
DC是一组不同类型的造血细胞,在先天免疫和后天免疫系统中发挥重要作用。3-5它们是有效的抗原感应和抗原呈递细胞(专职性抗原提呈细胞APC),能够独特地启动针对外来抗原的初次免疫应答,同时保护对自体抗原的耐受性。6 DC指导免疫应答的特异性、大小和极性。
树突状细胞成熟
未成熟的DC起源于骨髓中的祖细胞,并迁移到全身几乎所有的淋巴组织和非淋巴组织,包括皮肤、肺和肠道。7,8多种转录因子、信号分子、生长因子、细胞因子、趋化因子和粘附受体参与了普通DC祖细胞到成熟DC的分化途径。3,9,10此外,通过多种表面模式识别受体(PRR),包括Toll样受体(TLR)和C型凝集素受体(CLR),未成熟DC识别其周围环境中的损伤相关分子模式(DAMP)或病原体相关分子模式(PAMP),从而接收和处理进一步成熟信号。3,12这种对受损细胞或病原体的感知使DC能够实现其前哨功能,以维持机体的完整性。
成熟中DC根据微环境信号改变其表面趋化因子受体和粘附分子的情况,并响应趋化信号进入次级淋巴器官。在淋巴组织内,未成熟驻留或传入非驻留DC可以进一步刺激分化为成熟的功能性DC。成熟DC具有高级功能来处理自身MHC抗原,并将其呈递给初始CD4+或CD8+T细胞。这导致针对外来抗原的初次免疫应答的启动,或针对自体抗原的潜在T细胞反应性的下调。成熟DC可以增加肽负载主要组织相容性复合体(MHC)抗原、共刺激(或共抑制)受体和配体(例如CD80和CD86)的表面表达,以及释放细胞因子(例如IL-6、IL-12p70或干扰素),从而刺激初始T细胞。12,13细胞可以进一步调节成熟DC的性质。应答的T细胞可相互调节DC,例如通过CD40-CD40L相互作用或通过T细胞衍生的细胞因子(如IL-4或IFN-γ)进行。通过这种方式,T细胞可以另外指导专职性抗原提呈细胞,这可以促进不同类型的T细胞依赖性免疫或耐受。

树突状细胞多功能作用
DC不仅是免疫应答的有效启动子,而且在决定随后发生的免疫应答的类型、大小和持续时间方面也起着重要的调节作用。1,4,10,11 DC通过差异表达其细胞表面配体和受体,以及分泌不同细胞因子、趋化因子和炎症介质,从而实现该作用。例如,释放IL-12p70的DC可优先促进1型CD4+辅助性T细胞(Th1)或细胞溶解性CD8+T细胞。其他类型DC可促进T细胞依赖性体液或细胞介导的免疫应答,为Th2、Th9、Th17、Th22、卵泡辅助性T细胞(Tfh)或调节性T细胞(Treg)的特征。究竟是哪种DC协调了这些类型的T细胞依赖性免疫应答,以及如何调节,这仍然未知,需要深入研究。
某些研究认为DC的成熟度很关键,而另一些研究则认为病原体类型或相关组织部位的重要影响很关键。这些都是需要仔细研究的关键参数。事实可能介于两者之间,因为DC通路中存在着很大程度的功能可塑性。15,16,17 DC在先天免疫和后天免疫之间的重要联系作用也越来越受到重视。DC在应答危险信号中成熟,从而能够诱导生产性T细胞应答。它们还能激活巨噬细胞、自然杀伤(NK)细胞、自然杀伤T细胞(NKT细胞)、粒细胞和肥大细胞,从而触发对入侵致病源的自然应答。15浆细胞样DC(pDC)是干扰素的主要来源,在应答某些病毒时迅速分泌,9这是DC在先天和后天免疫应答中发挥多功能作用的重要例子。
树突状细胞异质性
多种类型的前体、未成熟和成熟的DC(例如,朗格汉斯细胞、真皮或间质DC、血液DC)在起源、形态、定位、成熟状态、表型和功能10,15方面存在差异,已有描述。尽管人类和小鼠的细胞表面表型存在差异,但在人类和小鼠模型系统中已经描述了两种普遍接受的DC类型,它们似乎代表不同的谱系:浆细胞样DC(pDC)和髓样DC(mDC),也称为经典或常规树DC(cDC)。1 pDC具有产生干扰素的巨大能力,但可能不像mDC那样有效地呈递抗原。1,3,人类pDC的特征在于其共表达CD123和CD304,而小pDC表达CD45R/B220和Ly-6C。1,9,10在人类和小鼠中,有两大类mDC被进一步分类,它们由干扰素调节因子4(IRF4+DC)或干扰素调节因子8(IRF-8+DC)的选择性表达来确定。11人类IRF4+DC特征性地表达CD1c,而小鼠对应物表达CD4(淋巴驻留DC)或CD11b(迁移DC)。两个物种的IRF4+ DC共表达CD172a/Sirp-α,能有效地将抗原呈递给初始CD4+T细胞。人类IRF8+ DC基本表达CD141,而小鼠等同物表达CD8a(淋巴驻留DC)或CD103(迁移DC),其所有亚群均表达XCR1趋化因子受体CD370/Clec9a,并且能够将抗原呈递给CD4+T细胞和CD8+T细胞。人类和小鼠朗格汉斯细胞(LC)同样共同表达几种不同的标记物,包括CD207/Langerin、CD326/EpCAM和CD324/E-Cadherin。3,10两个物种的真皮和肠道中的DC亚群也已描述。3人类和小鼠DC对应物的总结见下表。
人类DC亚群及小鼠DC对应物的功能性
| 人类DC亚群 | 小鼠DC对应物 | 比例 | 定位 | 刺激产生细胞因子* |
|---|---|---|---|---|
| pDC | pDC | ~1% 的外周血单核细胞(PBMC) | 人血 淋巴结 T细胞区 扁桃体 |
IFN-I+, IFN-III (IFN-λ)+ IL-6+, IL-8+ IP-10 (CXCL10)+ TNF+ |
| CD1c+ DCs | CD4+ or CD11b+ DCs | ~1% PBMCs | 人血 非淋巴组织: 皮肤、肝脏、肺和肠道 淋巴组织: 脾脏、淋巴结 |
IL-1β+, IL-6+, IL-8+
IL-10+, IL-12+ IL-23+ TNF+ IL-15+ (皮肤) |
| CD141+ | CD8+ or CD103+ DCs | 0.03% PBMCs CD8+ DCs: 20–40% 的小鼠脾脏和淋巴结 cDC | 人类淋巴结、扁桃体、脾脏、骨髓 人类非淋巴组织: 皮肤、肺、肝脏、肠道 CD8+ DCs: 小鼠淋巴组织 |
IFN-I+, IFN-III (IFN-λ)+
IL-12+ (小鼠) CXCL-10 (IP-10)+ TNF+** |
| LCs (朗格汉斯细胞) | LCs (朗格汉斯细胞) | 3–5% 表皮细胞 | 人类复层鳞状上皮, 引流淋巴结 | IL-15+ |
| 炎性DC | 炎性DC | 炎性位点 | IL-1β+, IL-6+
IL-10+, IL-12+, IL-23+ TNF+ |
*细胞因子的产生可能因所用刺激剂、刺激条件或细胞生理状态而异。
**TNF通常不是由人类CD141+DC在应答TLR8刺激下产生的。
另一类DC,即炎性DC,可能由单核细胞产生,这些单核细胞可能受环境刺激的驱动,具有DC的特性和功能。3显然,不同DC亚群的刺激性物种间差异以及某些功能相关分子的相似性正在被研究,包括它们的TLR、CLR、CD1分子、趋化因子受体的表达情况及其细胞因子分泌模式。3由于DC亚群和成熟阶段等多种因素共同影响T细胞应答,进一步研究DC生物学在生理和病理条件下复杂性时,详细表型分析结合功能研究将是一种有效途径。
参考
- Palucka K and Banchereau J. Cancer immunotherapy via dendritic cells. Nat Rev Cancer. 2012;12(4):265-277. doi: 10.1038/nrc3258
- Janco JMT, Lamichhane P, Karyampudi L, Knutson K. Tumor-infiltrating dendritic cells in cancer pathogenesis. J Immunol. 2015;194(7):2985-2991. doi: 10.4049/jimmunol.1403134
- Merad M, Sathe P, Helft J, Miller J, Mortha A. The dendritic cell lineage: ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Annu Rev Immunol. 2013;31:563-604. doi: 10.1146/annurev-immunol-020711-074950
- Schraml BU, Reis e Sousa C. Defining dendritic cells. Curr Opin Immunol. 2015;32:13-20. doi: 10.1016/j.coi.2014.11.001
- Collin M, McGovern N, Haniffa M. Human dendritic cell subsets. Immunology. 2013;140(1):22-30. doi: 10.1111/imm.12117
- O'Keeffe M, Mok WH, Radford KJ. Human dendritic cell subsets and function in health and disease. Cell Mol Life Sci. 2015;72(22):4309-4325. doi: 10.1007/s00018-015-2005-0
- Apostolopoulos V, Thalhammer T, Tzakos AG, Stojanovska L. Targeting antigens to dendritic cell receptors for vaccine development. J Drug Deliv. 2013;2013:869718. doi: 10.1155/2013/869718
- Cohn L, Delamarre L. Dendritic cell-targeted vaccines. Front Immunol. 2014;5:255. doi: 10.3389/fimmu.2014.00255
- Delamarre L, Mellman I. Harnessing dendritic cells for immunotherapy. Sem Immunol. 2011;23(1):2-11. doi: 10.1016/j.smim.2011.02.001
- Breton G, Lee J, Liu K, Nussenzweig MC. Defining human dendritic cell progenitors by multiparametric flow cytometry. Nat Protoc.2015;10(9):1407-1422. doi: 10.1038/nprot.2015.092
- Poltorak MP, Schraml BU. Fate mapping of dendritic cells. Front Immunol. 2015;6:199. doi: 10.3389/fimmu.2015.00199
- Swiecki M, Colonna M. The multifaceted biology of plasmacytoid dendritic cells. Nat Rev Immunol. 2015;15(8):471-485. doi: 10.1038/nri3865
- Murphy TL, Grajales-Reyes GE, Wu X, et al. Transcriptional control of dendritic cell development. Annu Rev Immunol. 2016;34:93-119. doi: 10.1146/annurev-immunol-032713-120204
- Durand M, Segura E. The known unknowns of the human dendritic cell network. Front Immunol. 2015;6:129. doi: 10.3389/fimmu.2015.00129
- Dutertre CA, Wang LF, Ginhoux F. Aligning bona fide dendritic cell populations across species. Cell Immunol. 2014;291(1-2):3-10. doi: 10.1016/j.cellimm.2014.08.006
- Schlitzer A, Ginhoux F. Organization of the mouse and human DC network. Curr Opin Immunol. 2014;26:90-99. doi: 10.1016/j.coi.2013.11.002
- Reis e Sousa C. Activation of dendritic cells: translating innate into adaptive immunity. Curr Opin Immunol. 2004;16(1):21-25. doi: 10.1016/j.coi.2003.11.007.
人类树突状细胞
通过流式细胞仪,可以用多种表面标记物和细胞内标记物来表征人类树突状细胞(DC)亚群。
以下外周血例子显示了基于可用BD OptiBuild™定制试剂的DC亚群识别
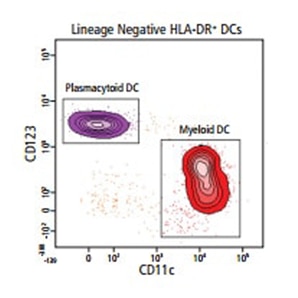

各种人类DC亚群表型特征的关键标记物见下表;细胞内标记物用粗体表示。BD Life Sciences拥有丰富的抗体试剂组合,以多种形式满足多数特异性,从而保证面板设计和下游分析的灵活性。
所有人类 DC 亚群被鉴定为谱系– (CD3– CD19– CD20– CD56– CD141) 和 CD45+ MHCII (HLA-DR)+ CD11c+/-2.
| DC 亚群 | 关键标记物 | |||
|---|---|---|---|---|
| 主要标记物 | 附加阳性标记物 | 附加阴性标记物 | 转录因子 | |
| 浆细胞样 DC | CD123 (IL-3Rα)高 CD303 (BDCA2/CLEC4C)+ CD304 (Neuropilin-1/BDCA4)+ CD85g (ILT7)+ CD11c低 CD11b–, CD14– |
CD2±, CD4+, CD45RA+ CD141 (BDCA3)低 CD272 (BTLA)+ CD366 (TIM-3)± CD367 (DCIR/CLEC4A)+ CD371 (CLEC12A)± TLR7高, TLR9高 FLT3 (CD135)+ GM-CSFR (CD116)+ CCR5+, CXCR4+ |
CD1a–, CD1c (BDCA1)– CD16 (FcγRIII)– CD172a (Sirp-α)– CD207 (Langerin)– CD324 (E-Cadherin)– CD326 (EpCAM)– CD369 (Dectin-1/CLEC7A)– CD370 (CLEC9A/DNGR1)– CLEC6A (Dectin-2)– XCR1– |
IRF7+, IRF8+ SpiB+ |
| CD1c+ 髓样 DC |
CD1c (BDCA1)+ CD172a (Sirp-α)+ CLEC6A (Dectin-2)+CD11b+/低, CD11c+ CX3CR1+CD14低/– |
CD4+, CD13+, CD26低, CD33+ CD45RO+, CD141 (BDCA3)± CD272(BTLA)+ CD366 (TIM-3)+ CD367 (DCIR/CLEC4A)+ CD369 (Dectin-1/CLEC7A)+ CD371 (CLEC12A)+ CD1a+ [皮肤和肠道] CD141(BDCA3)+ [肠道] TLR3低, TLR4低, TLR8+, TLR10低 FLT3 (CD135)+, GM-CSFR (CD116)+ |
CD1a–, CD16 (FcγRIII)– CD123 (IL-3Rα)– CD207 (Langerin)– CD304 (Neuropilin-1/BDCA4)–, CD324 (E-Cadherin)– CD326 (EpCAM)– CD370 (CLEC9A/DNGR1)– ESAM– XCR1– | IRF4+ |
| CD141+ 髓样 DCs |
CD141 (BDCA3)高 CD370 (CLEC9A/DNGR1)+ NECL2 (CADM1)+ CD11c+/低 CD14– |
CD4+, CD11b低 CD26+, CD162高 CD205 (DEC-205)高 CD272 (BTLA)高 CD367 (DCIR/CLEC4A)+ CD366 (TIM-3)+ CD369 (Dectin-1/CLEC7A)+ CD371 (CLEC12A)+ TLR3+, TLR8+ FLT3 (CD135)+ GM-CSFR (CD116)+ XCR1+ |
CD1a−, CD1c (BDCA1)−, CD16 (FcγRIII)− CD172a (Sirp–α)− CD207 (Langerin)− CD304 (Neuropilin-1/BDCA4)− CD324 (E-Cadherin)− CD326 (EpCAM)− |
IRF8+ BATF3+ |
| 朗格汉斯细胞 | CD207 (Langerin)+ CD324 (E-Cadherin)+ CD326 (EpCAM)+ CD11blow, CD11c+ CD14− |
CD1a高, CD1c (BDCA1)+ CD36+ CD172a (Sirp-α)+ CD369 (Dectin-1/CLEC7A)+, CD371 (CLEC12A)+ CLEC6A (Dectin-2)+ TLR1+, TLR2+, TLR3低, TLR6+ |
CD304 (Neuropilin-1/BDCA4)– CD367 (DCIR/CLEC4A)– XCR1– |
|
| CD1a+ 真皮 DC |
CD1a+ CD64 (FcγRI)+ CD366 (TIM-3)+ CD11b+, CD11c+ CD14− |
CD1c (BDCA1)+ CD172a (Sirp-α)+ CD367 (DCIR/CLEC4A)+ CD369 (Dectin-1/CLEC7A)+ CLEC6A (Dectin-2)+ TLR1–3+, TLR6+, TLR7+, TLR10+ |
CD207 (Langerin)− CD209 (DC-SIGN/CLEC4L)− CD324 (E-Cadherin)− CD326 (EpCAM)− |
|
| CD14a+ 真皮 DC |
CD14+ CD209 (DC-SIGN/CLEC4L)+ CD11b+, CD11c+ |
CD1c (BDCA1)+ CD172a (Sirp-α)+ CD367 (DCIR/CLEC4A)+ CD369 (Dectin-1/CLEC7A)+ CLEC6A (Dectin-2)+ TLR1–3+, TLR6, TLR7+ CSF–1R (CD115)+ |
CD1a− CD207 (Langerin)− CD324 (E-Cadherin)− CD326 (EpCAM)− CD366 (TIM-3)− |
|
| 炎性 DC (单核细胞源 DCs) |
CD16 (FcγRIII)+ CD64 (FcγRI)+ CD1a+ CD1c (BDCA1)+ CD11b+, CD11c+ CD14± |
CD172a (Sirp-α)+ CD206 (MR/CLECL13D)+ CD209 (DC-SIGN/CLEC4L)+ CD367 (DCIR/CLEC4A)+ CD369 (Dectin-1/CLEC7A)+ CD371 (CLEC12A)+ CLEC6A (Dectin-2)+ TLR3低, TLR4+, TLR7低, TLR8+ CCR2 (CD192)+ |
CD207 (Langerin)− CD324 (E-Cadherin)− CD326 (EpCAM)− |
1CD14 在所有DC亚群中均为阴性或低表达,CD14+真皮DC和炎性DC除外。
2CD11c 在所有DC亚群中均为阳性,但浆细胞样DC除外,其表达为低表达或阴性。
TLR7、 8 和 9 是内体的,需要细胞内染色。
小鼠树突状细胞
通过流式细胞仪,可以用多种表面标记物和细胞内标记物来表征小鼠树突状细胞(DC)亚群。
以下小鼠脾脏例子显示了基于可用BD OptiBuild™定制试剂的DC亚群识别
小鼠树突状细胞亚群的免疫分型。小鼠脾脏中三个主要的DC亚群(髓样、淋巴和浆细胞样)的10色免疫表型特征。用酶消化BALB/c小鼠脾脏,用抗体合剂(包括OptiBuild™试剂SirpαBV650和Clec12A BV786)染色,用BD FACSCelesta™流式细胞仪分析,细胞仪采用蓝/紫/红(BVR)激光配置。首先在CD3, CD19 and 7-AAD阴性细胞上设门(未显示)。A-D. 设门策略:I-A/I-E高CD11c高 常规树突状细胞(DC)进一步分化为CD11b+mDC和CD8+lDC。I-A/I-E低CD11c低 B220+Gr1+细胞被视为pDC。结果:在mDC(E、F、G),lDC(H、I、J)和pDC(K、L、M)亚群中,分别进一步分析额外标记物CD4、CD172a(Sirpα)和CD371(Clec12A)的差异表达。门按荧光扣除(FMO)对照设置。
各种小鼠DC亚群表型特征的关键标记物见下表;细胞内标记物用粗体表示。BD Life Sciences拥有丰富的抗体试剂组合,以多种形式满足多数特异性,从而保证面板设计和下游分析的灵活性。
小鼠DC标记物被鉴定为谱系– (CD3– CD19– CD49b– or NK1.1– CD14–)和CD45+ MHCII+ CD11c+。

DC亚群 | 关键标记物 | |||
主要标记物 | 附加阳性标记物 | 附加阴性标记物 | 转录因子 | |
浆细胞样DC (pDC) | CD45R (B220)+ CD317 (BST-2)+ Ly-6C+ Siglec-H+ CD11c低 CD14– | CD4+, CD26+ CD172a (Sirp-α)+ CD209a (DC-SIGN)+ CD272 (BTLA)+ CD370 (Clec9a/DNGR1)+ Gr1 (Ly-6C 和 Ly-6G)+ FLT3 (CD135)+ TLR7高, TLR9高 | CD11b– CD24– CD36–, CD64 (FcγRI)–, CD103– CD205 (DEC-205)– CD207 (Langerin)– CD326 (EpCAM)– DCIR2 (Clec4a4/33D1)– F4/80– CX3CR1–, XCR1– | IRF7+, IRF8+ BATF3高 SpiB+
Zbtb46– |
CD4+CD11b+ 淋巴驻留DC | CD4+ CD11b+ CD11c高 CD8a– CD14– | CD24+, CD26+ CD172a (Sirp-α)+ CD205 (DEC-205)+ CD209a (DC-SIGN)+ CD272 (BTLA)低 DCIR2 (Clec4a4/33D1)+ ESAM+ F4/80+ FLT3 (CD135)+ TLR5+, TLR7+, TLR9+ CX3CR1+ | CD36– CD45R (B220)– CD64 (FcγRI)–, CD103– CD207 (Langerin)– CD326 (EpCAM)– CD370 (Clec9a/DNGR1)– Ly6C– XCR1– | IRF4+ Zbtb46+ BATF3高 |
CD4–CD11b+ 常规迁移DC | CD11b+ CD11c+ CD4– CD8a– CD14– | CD24±, CD26+ CD64 (FcγRI)+ CD172a (Sirp-α)+ CD209a (DC-SIGN)± CD272 (BTLA)低 DCIR2 (Clec4a4/33D1)+ F4/80+ Ly-6C± FLT3 (CD135)+ TLR5+, TLR7+, TLR9+ CX3CR1+ | CD36– CD45R (B220)–, CD103– CD207 (Langerin)– CD326 (EpCAM)– CD370 (Clec9a/DNGR1)– XCR1– | IRF4+ IRF2+ Zbtb46+ BATF3高 |
CD8a+ 常规 淋巴驻留DC | CD8a+ CD11c高 CD4– CD11b– CD14– | CD1d1+, CD24+, CD26+ CD36+, CD103± CD205 (DEC-205)+ CD207 (Langerin)± CD272 (BTLA) 高 CD370 (Clec9a/DNGR1)+ NECL2 (CADM1)+ FLT3 (CD135)+ TLR3+, TLR4+, TLR11+ XCR1+, CX3CR1± | CD45R (B220)– CD64 (FcγRI)– CD172a (Sirp-α)– CD209a (DC-SIGN)– CD326 (EpCAM)– DCIR2 (Clec4a4/33D1)– F4/80– Ly-6C– | IRF8+ BATF3+ Zbtb46+ |
CD103+ 常规 迁移DC | CD103+ CD11c高 CD4– CD8– CD11b– CD14– | CD1d1+, CD24+ CD26+, CD36+ CD205 (DEC-205)+ CD207 (Langerin)+ CD272 (BTLA) 高 CD370 (Clec9a/DNGR1)+ NECL2 (CADM1/CD317)+ FLT3 (CD135)+ TLR3+, TLR4+, TLR11+ XCR1+ | CD45R (B220)– CD64 (FcγRI)– CD172a (Sirp-α)– CD209a (DC-SIGN)– CD326 (EpCAM)– DCIR2 (Clec4a4/33D1)– F4/80– Ly-6C– CX3CR1– | IRF8+ BATF3+ Zbtb46+ |
朗格汉斯细胞 | CD207 (Langerin)+ CD324 (E-Cadherin)+ CD326 (EpCAM)+ CD11b+ CD11c+ CD14– | CD24+ CD172a (Sirp-α)+ CD205 (DEC-205)+ F4/80+ TLR3+, TLR11+ | CD8a–, CD26–, CD36– CD45R (B220)–, CD103– CD209a (DC-SIGN)– CD370 (Clec9a/DNGR1)– DCIR2 (Clec4a4, 33D1)– Ly-6C– CX3CR1–, XCR1– |
|
CD207+ 真皮DCs | CD207 (Langerin)+ CD11b低 CD11c+ | CD103± | CD45R (B220)– CD172a (Sirp-α)– CD326 (EpCAM)– |
|
CD207– 真皮DC | CD207 (Langerin)– CD11b± CD11c+ | CD172a (Sirp-α)+ | CD45R (B220)– CD103– CD326 (EpCAM)– |
|
炎性DC (单核细胞源DC) | CD64 (FcγRI)+ CD11b+ CD11c+ CD14– | CD209a (DC-SIGN)+ CD272 (BTLA)低 Ly-6C+ TLR1-6, TLR7-8+, TLR10+ | CD8a–, CD45R (B220)–, CD103– CD172a (Sirp-α)– CD207 (Langerin)– CD326 (EpCAM)– |
|
树突状细胞的分离
某些研究方法需要使用树突状细胞(DC)的分离群体,进行进一步的特性、培养或功能研究。分离这些非常罕见的细胞类型(仅占血液单核细胞的0.1–1%)在以往一直很困难。3流式细胞仪和磁珠分离技术的进步使DC分离变得更容易。基于流式细胞仪的细胞分选法可以分离单细胞,以进行进一步分析。
树突状细胞群体
分离的DC群体通常用于功能分析,例如与T细胞共培养,或用于其他下游应用。BD Biosciences提供了成功分离人类DC和小鼠DC的方法。
BD IMag™细胞分离系统基于一种简单高效的直接磁铁技术。例如,从一系列BD IMag™颗粒格式的细胞表面结合抗体中,研究人员正选择或负选择DC是可行的。易于使用的人类和小鼠DC富集子集,在几个短步骤内负选择非DC群体,也是可用的。
分离这些极为罕见的细胞,流式细胞仪分选是一种非常有效的方法,因为它提供了高纯度和细胞回收,非常适合分离由多标记物表型定义的细胞。许多研究人员依靠BD-FACSAria™ 细胞分选机分离特定DC群体。5
从人类PBMC中分离pDC和mDC
基于Lin– HLA-DR+ CD123+ CD11c– (pDCs)和Lin– HLA–DR+ CD123– CD11c+(mDCs)表型,可以从人外周血中分离出浆细胞样DC(pDC)和髓样DC(mDC)。在以下示例中,通过BD IMag™人类树突状细胞富集装置(编号558420),对外周血单核细胞(PBMC),执行预富集步骤,负选择红细胞、血小板和外周白细胞(不是DC),从而提供DC富集样品用于细胞表面染色和分选。
用谱系合剂FITC、CD123 BV421、HLA-DR PE和CD11c APC进行细胞染色。用活性染料FVS780排除死细胞。用BD FACSAria™流式细胞仪分选细胞。BD IMag™基于颗粒富集结合流式细胞仪分选,这是一种方便省时的工作流程,产生高纯度的DC群体,这些群体通常很少在人体血液中发现。
BD应用程序专家可提供现场或电话咨询,以支持您的分选程序。有关更多信息,请访问我们的支持页面。
BD流式细胞仪平台,如BD FACSMelody™、BD FACSAria™ III、BD FACSAria™融合细胞分选机,支持将单细胞分选纳入不同类型的平板。自动沉积单元使单细胞分选成为可能,并且当与索引分选功能结合时,该效用得到增强,索引分选功能记录流式细胞仪数据(例如,标记物表型)和每个分选事件的分选位置(分选收集设备的X坐标和Y坐标)。这样,分选后分析的结果(可能包括DNA或RNA序列分析),可以精确地追溯到特定细胞的流动特征。

细胞因子检测
通过测量树突状细胞(DC)分泌的细胞因子的类型和数量,可以深入了解DC应答的性质和程度。多重分析越来越多地用于测定分泌因子,如细胞因子和趋化因子。
与传统的酶联免疫吸附试验(ELISA)相比,荧光检测和多重测量有广泛动态范围,使得样品量更小、样品稀释更少,并且基本需要更短时间来确定未知值。
BD™CBA组合可以用于测量分析多种可溶性因子,包括分泌的蛋白质(例如细胞因子或趋化因子)、脱落或释放的细胞表面标记物(例如sCD14)以及细胞信号分子(例如细胞裂解物内的磷蛋白)。
细胞内细胞因子的检测
通过使用蛋白转运抑制剂来阻断分泌,可以在产生细胞因子的细胞中检测细胞因子。因此,有可能区分被激活细胞群体产生细胞因子的详情:是少数细胞产生大量细胞因子;还是大量细胞参与,每个细胞产生少量细胞因子。

转录因子和其他细胞内蛋白
从早期发育到成熟DC的效应器功能,对DC生理学的分析,揭示了与发育状态、细胞激活或其他功能特性相关的细胞内变化。
BD提供了能够分析这些细胞内分子的解决方案,以支持研究人员破译调控DC生物学的相互关联的通路。
转录因子的检测
BD提供几种DC相关的经流式细胞仪验证的转录因子抗体。例如,在小鼠中,Zbtb46在常规DC及其前体中表达,但在pDC中不表达。而pDC的发育和功能需要Spi-B。2

磷蛋白的检测
BD Phosflow™抗体是单克隆磷酸表位特异性抗体,经验证可用于流式细胞术检测。对于大多数BD Phosflow™抗体,推荐通透性缓冲液是BD Phosflow™ Perm Buffer III,但是也可以使用替代的通透性缓冲液,可以用于细胞表面染色进行亚群分析。
许多BD Phosflow™抗体特异性可用于分析参与DC发育和激活的信号通路,例如对pDC发育有不同影响的pSTAT3和pSTAT5试剂。BD Phosflow™系统已用于pDC病毒感染后体外检测磷蛋白(如pSyk)3,广泛应用于先天免疫和后天免疫细胞信号学研究。

免疫荧光和免疫组织化学
DC形态与亚细胞定位
使用荧光成像单克隆抗体可以识别组织内的DC,并了解DC的体外生物学机制。
BD单克隆抗体直接与新的明亮的光稳定荧光染料结合,如BD Horizon Brilliant Violet™ 421 (BV421)和BD Horizon™ BV480,使多色成像超越了传统的三色到四色显微镜技术。
")
参考
- Merad M, Sathe P, Helft J, Miller J, Mortha A. The dendritic cell lineage: ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Annu Rev Immunol. 2013;31:563-604. doi: 10.1146/annurev-immunol-020711-074950
- Collin M, McGovern N, Haniffa M. Human dendritic cell subsets. Immunology. 2013;140(1):22-30. doi: 10.1111/imm.12117
- Schraml BU, Reis e Sousa C. Defining dendritic cells. Curr Opin Immunol. 2015;32:13-20. doi: 10.1016/j.coi.2014.11.001
仅供研究使用,不用于诊疗程序。
Alexa Fluor是Life Technologies Corporation公司的商标。