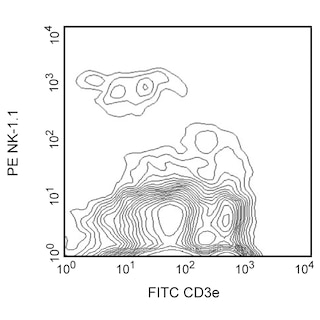
The 4D11 antibody specifically recognizes Ly-49G2 (also known as LGL-1), an inhibitory receptor which is expressed on subsets of natural killer (NK) cells and DX5-positive T lymphocytes (NK-T cells) in all strains tested (e.g., AKR/N, BALB/c, C3H/HeJ, C57BL/6, CBA/J, DBA/2, SJL, 129) and on a population of memory CD8+ T lymphocytes in C57BL/6 mice. Cross-reaction of 4D11 antibody to Ly-49A[B6], Ly-49A[BALB], and Ly-49T[129/J] inhibitory receptors and Ly-49L[CBA/J] activating receptor has been reported. The proportion of NK-T cells expressing Ly-49A and Ly-49G2 is higher (2-5 fold) in thymus than in liver (immature and mature NK-T cells, respectively), and there is evidence that down-regulation of Ly-49 receptor expression is necessary for normal NK-T-cell development to occur. Most NK cells express a single allele of Ly-49A and/or Ly-49G2, although occasionally they may express more than one allele. The Ly-49 family of NK-cell receptors, members of the C-type lectin superfamily, are disulfide-linked type-II transmembrane protein homodimers with extracellular carbohydrate-recognition domains, which bind to MHC class I alloantigens. The Ly-49 family members are expressed independently, such that an individual NK or T cell may display more than one class of Ly-49 receptor homodimers. Binding of Ly-49G[B6]-expressing transfectants to H-2Dd+/H-2Ld+ ConA blasts has been demonstrated, and H-2D[d]-expressing target cells inhibit the lytic activity of Ly-49G2-expressing NK cells. The levels of the Ly-49 inhibitory receptors are down-regulated by their ligands in vivo, and the various levels of expression of a Ly-49 inhibitory receptor may affect the specificity of NK cells. Ly-49G2[+] NK cells are able to lyse target tumor cells expressing H-2[a] and H-2[b] MHC class I antigens in vitro, and they mediate allogeneic and hybrid resistance to H-2[b] bone marrow transplantation. The Ly-49A[BALB] and Ly-49A[B6] alloantigens bind to MHC class I antigens of the d and k haplotypes, and Ly-49A[+] IL-2-activated NK cells are unable to lyse target cells expressing H-2[d] and H-2[k]. In vitro studies suggest that the Ly-49G2 and Ly-49A receptors mediate negative regulation of NK-cell cytolytic activity via tyrosine phosphorylation of their ITIMs (Immunoreceptor Tyrosine-based Inhibitory Motifs). Ly-49T[129/J] has a unique ITIM sequence, and Ly-49T-transfected 293T (human kidney epithelial) cells do not bind soluble tetramers of any tested H-2 alloantigen (D[b], D[d], D[k], K[b], K[d], K[k], L[d]).



.png?imwidth=320)