Overview
Killer cell lectin-like receptor G1 (KLRG1) is an immune checkpoint receptor encoded by the KLRG1 gene. It is an inhibitory lectin-like type II transmembrane receptor containing a cytoplasmic immunoreceptor tyrosine-based inhibitory motif (ITIM).1 KLRG1 is primarily composed of a homodimeric molecule consisting of two 30–38 kDa N-glycosylated subunits. BD carries several clones for the detection of KLRG1 in both human and mouse cells.
KLRG1 Biology
Function
E-, N- and R-cadherins are ligands of KLRG1; binding prevents Akt phosphorylation and increases expression of cell cycle inhibitors.2 Binding also recruits signaling molecules SHP-2 and SHIP-1, which in turn inhibit NK effector function and T cell proliferation. KLRG1 expression is correlated with reduction of both activated T lymphocyte proliferation and activated NK cell effector function.3 KLRG1 plays a role in leucocyte regulation in both the innate and adaptive immune systems. It is considered a senescent or differentiation cell marker. For example, KLRG1 can be used to define CD8 T cell subsets in acute infection4 and cancer5 to differentiate between short-lived effector (KLRG1high CD127low) and memory precursor (KLRG1low CD127high) CD8 T cells.6
KLRG1 as a Tool for Understanding Acute Immune Response
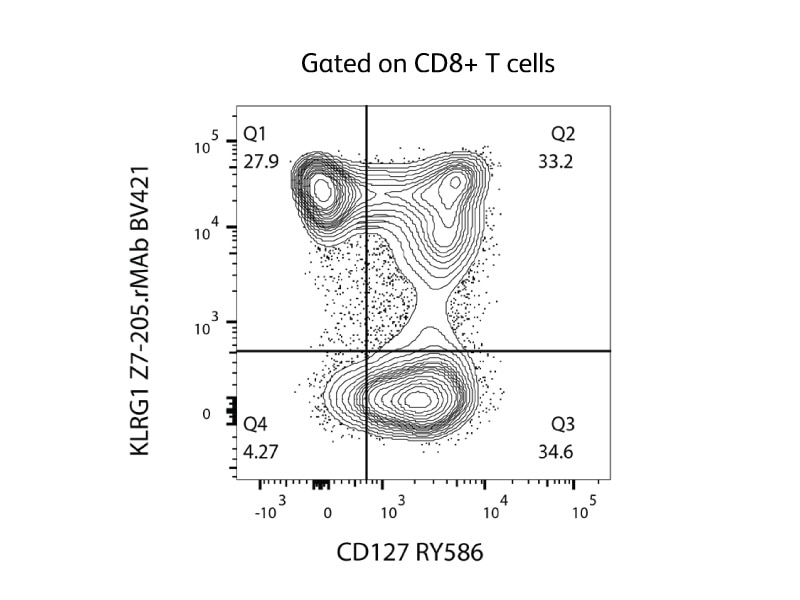
During acute infection, antigen-specific CD8 T cells expand and differentiate CD8 memory precursor effector cells (MPECs), which express KLRG1low CD127high, and short-lived effector cells (SLECs), which express KLRG1high CD127low.7 CD127, the receptor for interleukin 7, can dictate a cell’s longevity by increasing the expression of anti-apoptotic factors; therefore, MPECs express lower levels of CD127 than SLECs.8 The corresponding figure shows the detection of MPECs (Q3) and SLECs (Q1) on CD8 T cells using fluorescent-conjugated antibodies against KLRG1 and CD127.
Differentiation of CD8 T cells into either MPECs or SLECs is determined by several transcription factors, whose expression is influenced by the local cytokine milieu. High levels of inflammatory cytokines such as IL-12 and IFN increase expression of transcription factors including T-bet and Blimp-1, which promotes SLEC differentiation; conversely, low levels of inflammatory cytokines increases expression of transcription factors such as Id3 and Eomes, which leads to MDEC development.9-10 BD supports the detection of transcription factors with a variety of reagents, including fluorescent-conjugated antibodies and permeabilization systems.
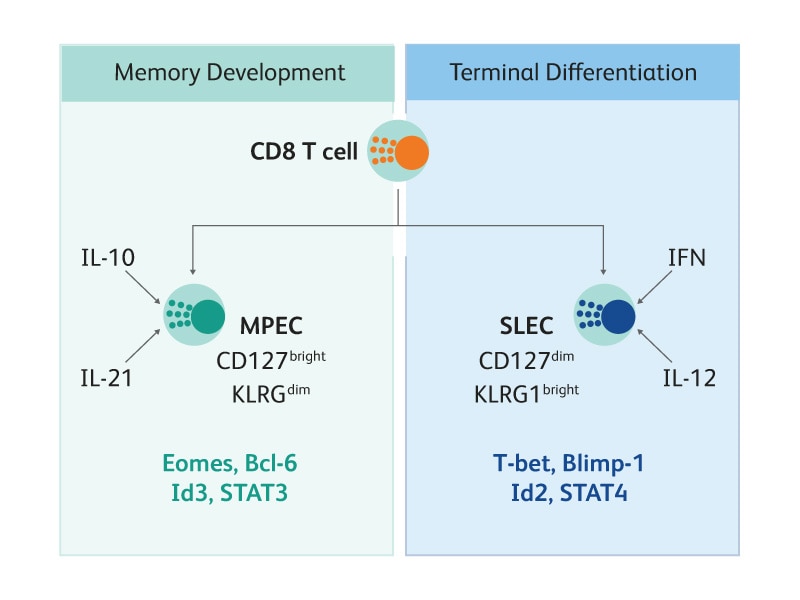
Differentiation of antigen-specific CD8 T cells into MPECs or SLECs is dependent on the inflammation factors present in the environment, which affects the expression of several transcription factors that drive development.11
KLRG1 in Disease
Studies report that KLRG1 is highly expressed by T cells in the tumor microenvironment making it a potential therapeutic target in immuno-oncology. 12, 13 Subsequently, anti-KLRG1 antibody treatment (alone or with anti-PD-1 therapy) in mouse cancer models can be effective for tumor regression and reducing metastasis.
Human KLRG1
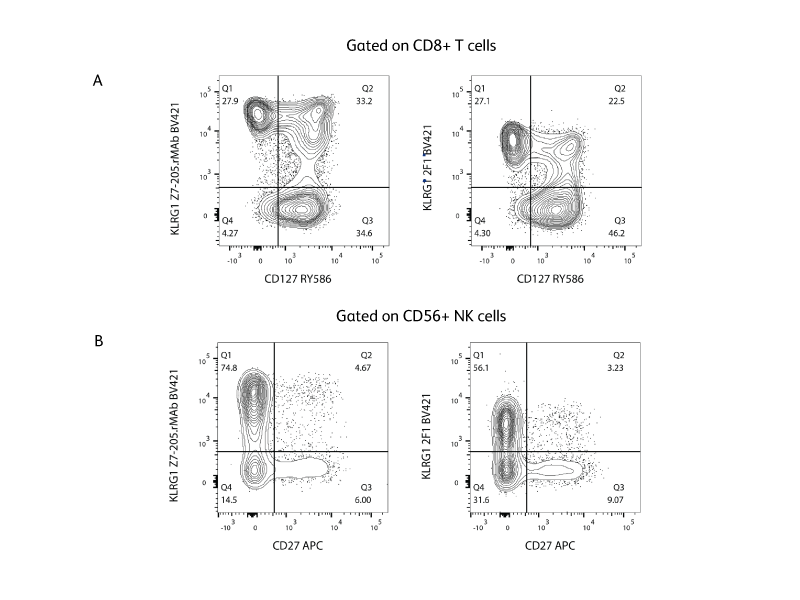
Human KLRG1 is expressed on a large subset of NK cells, lymphokine-activated killer (LAK) cells, adherent LAK (A-LAK) cells and subsets of CD8+ and CD4+ T lymphocytes but not mast cells.1 BD developed a novel recombinant monoclonal antibody (rMab) targeting human KLRG1, which can improve separation and may help distinguish novel immune cell subsets expressing KLRG1.
Mouse KLRG1
Mouse KLRG1 is the homolog of the rat mast cell function-associated antigen (MAFA), on all mouse strains tested (eg, AKR/J, BALB/c, C3H/HeN, C3H.SW, C57BL/6, DBA/1, SJL, 129/J). Unlike rat MAFA, which is expressed on mast cells, mouse KLRG1 is expressed on a large subset of NK cells, lymphokine-activated killer (LAK) cells, adherent LAK (A-LAK) cells, subsets of activated CD8+ T lymphocytes and small fractions of CD4+ and CD8+ T cells but not mast cells.2, 14 KLRG1 expression is reduced in MHC class I-deficient mice, although direct binding of KLRG1 to MHC class I antigens could not be detected.15
Clone 2F1 is specific against mouse KLRG1 and is available in multiple formats and kits including BD® AbSeq and BD Lyoplate™ Screening Panels. Though 2F1 can be cross-reactive against human KLRG1, we suggest using clone Z7-205.rMAb for detection of human KLRG1 due to its brighter staining.
Clone 2F1 was used for multiomic analysis in the characterization of the tumor response of immune cell Clone 2F1 was used for multiomic analysis in the characterization of the tumor response of immune cell populations in a mouse model for B-cell lymphoma. The immune population of splenocytes from a mouse model for B-cell lymphoma were analyzed and sorted into several clusters based on a 21-color cell sorting panel. Protein and mRNA expression analysis of these clusters were analyzed with a 34-plex BD® AbSeq Panel. The heat map below shows differential protein and mRNA analysis, including KLRG1, in the endogenous CD8 T cell cluster.
Read datasheet entitled “Leveraging the power of high-parameter cell sorting and single-cell multiomics to profile intratumoral immune cells in a model of B-cell lymphoma”.
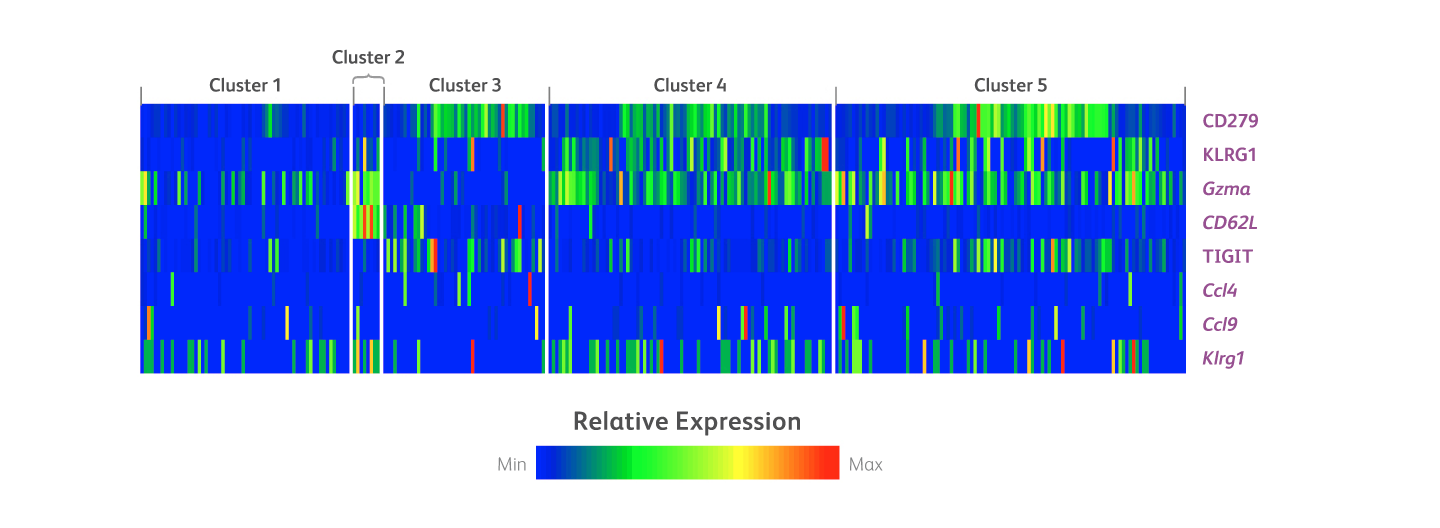
Heat map showing differential protein and mRNA analysis, including KLRG1, of clustered immune cell populations in a mouse model for B-cell lymphoma.
References
- Beyersdorf NB, Ding X, Karp K, Hanke T. Expression of inhibitory "killer cell lectin-like receptor G1" identifies unique subpopulations of effector and memory CD8T cells. Eur J Immunol. 2001; 31(12):3443-3452.
- Henson SM, Akbar AN. KLRG1--more than a marker for T cell senescence. Age (Dordr). 2009;31(4):285-291. doi:10.1007/s11357-009-9100-9
- Henson SM, Franzese O, Macaulay R, et al. KLRG1 signaling induces defective Akt (ser473) phosphorylation and proliferative dysfunction of highly differentiated CD8+ T cells. Blood. 2009;113(26):6619-6628. doi:10.1182/blood-2009-01-199588
- Wang D, Diao H, Getzler AJ, et al. The Transcription Factor Runx3 Establishes Chromatin Accessibility of cis-Regulatory Landscapes that Drive Memory Cytotoxic T Lymphocyte Formation. Immunity. 2018; 48(4):659-674.e6.
- Tata A, Dodard G, Fugère C, et al. Combination blockade of KLRG1 and PD-1 promotes immune control of local and disseminated cancers. Oncoimmunology. 2021;10(1):1933808. Published 2021 Jun 15. doi:10.1080/2162402X.2021.1933808
- Joshi NS, Cui W, Chandele A, et al. Inflammation directs memory precursor and short-lived effector CD8(+) T cell fates via the graded expression of T-bet transcription factor. Immunity. 2007;27(2):281-295. doi:10.1016/j.immuni.2007.07.010
- Joshi NS, Kaech SM. Effector CD8 T cell development: a balancing act between memory cell potential and terminal differentiation. J Immunol. 2008;180(3):1309-1315. doi:10.4049/jimmunol.180.3.1309
- Amsen D, Backer RA, Helbig C. Decisions on the road to memory. Adv Exp Med Biol. 2013;785:107-120. doi:10.1007/978-1-4614-6217-0_12
- Obar JJ, Jellison ER, Sheridan BS, et al. Pathogen-induced inflammatory environment controls effector and memory CD8+ T cell differentiation. J Immunol. 2011;187(10):4967-4978. doi:10.4049/jimmunol.1102335
- Plumlee CR, Obar JJ, Colpitts SL, et al. Early effector CD8 T cells display plasticity in populating the short-lived effector and memory-precursor pools following bacterial or viral infection. Scientific Reports. 2015;5(1). doi:10.1038/srep12264
- Joshi N, Cui W, Chandele A, et al. Inflammation Directs Memory Precursor and Short-Lived Effector CD8+ T Cell Fates via the Graded Expression of T-bet Transcription Factor. Immunity. 2007;27(2):281-295. doi:10.1016/j.immuni.2007.07.010
- Greenberg SA, Kong SW, Thompson E, Gulla SV. Co-inhibitory T cell receptor KLRG1: Human cancer expression and efficacy of neutralization in murine cancer models. Oncotarget. 2019;10(14):1399-1406. doi:10.18632/oncotarget.26659
- Akhmetzyanova I, Zelinskyy G, Littwitz-Salomon E, et al. CD137 Agonist Therapy Can Reprogram Regulatory T Cells into Cytotoxic CD 4+ T Cells with Antitumor Activity. J Immunol. 2016; 196(1):484-92.
- Hanke T, Corral L, Vance RE, Raulet DH. 2F1 antigen, the mouse homolog of the rat "mast cell function-associated antigen", is a lectin-like type II transmembrane receptor expressed by natural killer cells. Eur J Immunol. 1998; 28(12):4409-4417.
- Corral L, Hanke T, Vance RE, Cado D, Raulet DH. NK cell expression of the killer cell lectin-like receptor G1 (KLRG1), the mouse homolog of MAFA, is modulated by MHC class I molecules. Eur J Immunol. 2000; 30(3):920-930.