
Preparation And Storage
Product Notices
- Since applications vary, each investigator should titrate the reagent to obtain optimal results.
- An isotype control should be used at the same concentration as the antibody of interest.
- Warning: Some APC-Cy7 and PE-Cy7 conjugates show changes in their emission spectrum with prolonged exposure to formaldehyde. If you are unable to analyze fixed samples within four hours, we recommend that you use BD™ Stabilizing Fixative (Cat. No. 338036).
- This product is subject to proprietary rights of Amersham Biosciences Corp. and Carnegie Mellon University and made and sold under license from Amersham Biosciences Corp. This product is licensed for sale only for research. It is not licensed for any other use. If you require a commercial license to use this product and do not have one return this material, unopened to BD Biosciences, 10975 Torreyana Rd, San Diego, CA 92121 and any money paid for the material will be refunded.
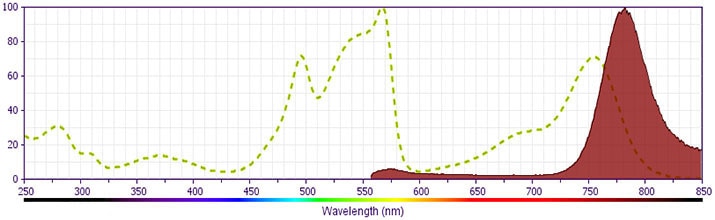
- PE-Cy7 is a tandem fluorochrome composed of R-phycoerythrin (PE), which is excited by 488-nm light and serves as an energy donor, coupled to the cyanine dye Cy7, which acts as an energy acceptor and fluoresces maximally at 780 nm. PE-Cy7 tandem fluorochrome emission is collected in a detector for fluorescence wavelengths of 750 nm and higher. Although every effort is made to minimize the lot-to-lot variation in the efficiency of the fluorochrome energy transfer, differences in the residual emission from PE may be observed. Therefore, we recommend that individual compensation controls be performed for every PE-Cy7 conjugate. PE-Cy7 is optimized for use with a single argon ion laser emitting 488-nm light, and there is no significant overlap between PE-Cy7 and FITC emission spectra. When using dual-laser cytometers, which may directly excite both PE and Cy7, we recommend the use of cross-beam compensation during data acquisition or software compensation during data analysis.
- Cy is a trademark of Amersham Biosciences Limited. This conjugated product is sold under license to the following patents: US Patent Nos. 5,486,616; 5,569,587; 5,569,766; 5,627,027.
- Caution: Sodium azide yields highly toxic hydrazoic acid under acidic conditions. Dilute azide compounds in running water before discarding to avoid accumulation of potentially explosive deposits in plumbing.
- Please observe the following precautions: Absorption of visible light can significantly alter the energy transfer occurring in any tandem fluorochrome conjugate; therefore, we recommend that special precautions be taken (such as wrapping vials, tubes, or racks in aluminum foil) to prevent exposure of conjugated reagents, including cells stained with those reagents, to room illumination.
- For fluorochrome spectra and suitable instrument settings, please refer to our Multicolor Flow Cytometry web page at www.bdbiosciences.com/colors.
- Please refer to www.bdbiosciences.com/us/s/resources for technical protocols.
Companion Products

.png?imwidth=320)
The MEL-14 monoclonal antibody specifically binds to CD62L (L-selectin), a 95 kDa (on neutrophils) or 74 kDa (on lymphocytes) receptor with lectin-like and Epidermal Growth Factor-like domains. In the mouse, L-selectin is detected on most thymocytes, with the highest levels of expression on an immunocompetent subset and a population of dividing progenitor cells, and on peripheral leukocytes, including subsets of B and T lymphocytes, neutrophils, monocytes, and eosinophils. This member of the selectin adhesion molecule family appears to be required for lymphocyte homing to peripheral lymph nodes and to contribute to neutrophil emigration at inflammatory sites. L-selectin is rapidly shed from lymphocytes and neutrophils upon cellular activation; metalloproteinases may mediate the release of CD62L ectodomains from the cell surface. The level of CD62L expression, along with other markers, distinguishes naive, effector, and memory T cells. L-selectin binds to sialytaed oligosaccharide determinants on high endothelial venules (HEV) in peripheral lymph nodes. In vitro studies have demonstrated that CD34, GlyCAM-1, and MAdCAM-1, all recognized by mAb MECA-79 (anti-mouse PNAd Carbohydrate Epitope, Cat. No. 553863), may be ligands for CD62L. MEL-14 mAb blocks in vitro binding of lymphocytes to peripheral lymph node HEV and inhibits in vivo lymphocyte extravasation into peripheral lymph nodes and late stages of leukocyte rolling.
Development References (20)
-
Cerwenka A, Carter LL, Reome JB, Swain SL, Dutton RW. In vivo persistence of CD8 polarized T cell subsets producing type 1 or type 2 cytokines. J Immunol. 1998; 161(1):97-105. (Biology). View Reference
-
Gallatin WM, Weissman IL, Butcher EC. A cell-surface molecule involved in organ-specific homing of lymphocytes. Nature. 1983; 304(5921):30-34. (Immunogen). View Reference
-
Iwabuchi K, Ohgama J, Ogasawara K, et al. Distribution of MEL-14+ cells in various lymphoid tissues. Immunobiology. 1991; 182(2):161-173. (Biology). View Reference
-
Jung TM, Gallatin WM, Weissman IL, Dailey MO. Down-regulation of homing receptors after T cell activation. J Immunol. 1988; 141(12):4110-4117. (Biology). View Reference
-
Kishimoto TK, Jutila MA, Berg EL, Butcher EC. Neutrophil Mac-1 and MEL-14 adhesion proteins inversely regulated by chemotactic factors. Science. 1989; 245(4923):1238-1241. (Biology). View Reference
-
Lanzavecchia A, Sallusto F. Dynamics of T lymphocyte responses: intermediates, effectors, and memory cells. Science. 2000; 290(5489):92-97. (Biology). View Reference
-
Lewinsohn DM, Bargatze RF, Butcher EC. Leukocyte-endothelial cell recognition: evidence of a common molecular mechanism shared by neutrophils, lymphocytes, and other leukocytes. J Immunol. 1987; 138(12):4313-4321. (Biology). View Reference
-
Ley K, Bullard DC, Arbones ML, et al. Sequential contribution of L- and P-selectin to leukocyte rolling in vivo. J Exp Med. 1995; 181(2):669-675. (Biology). View Reference
-
Mobley JL, Dailey MO. Regulation of adhesion molecule expression by CD8 T cells in vivo. I. Differential regulation of gp90MEL-14 (LECAM-1), Pgp-1, LFA-1, and VLA-4 alpha during the differentiation of cytotoxic T lymphocytes induced by allografts. J Immunol. 1992; 148(8):2348-2356. (Biology). View Reference
-
Peschon JJ, Slack JL, Reddy P, et al. An essential role for ectodomain shedding in mammalian development. Science. 1998; 282(5392):1281-1284. (Biology). View Reference
-
Pizcueta P, Luscinskas FW. Monoclonal antibody blockade of L-selectin inhibits mononuclear leukocyte recruitment to inflammatory sites in vivo. Am J Pathol. 1994; 145(2):461-469. (Biology). View Reference
-
Reichert RA, Jerabek L, Gallatin WM, Butcher EC, Weissman IL. Ontogeny of lymphocyte homing receptor expression in the mouse thymus. J Immunol. 1986; 136(10):3535-3542. (Biology). View Reference
-
Reichert RA, Weissman IL, Butcher EC. Dual immunofluorescence studies of cortisone-induced thymic involution: evidence for a major cortical component to cortisone-resistant thymocytes. J Immunol. 1986; 136(10):3529-3534. (Biology). View Reference
-
Reichert RA, Weissman IL, Butcher EC. Phenotypic analysis of thymocytes that express homing receptors for peripheral lymph nodes. J Immunol. 1986; 136(10):3521-3528. (Biology). View Reference
-
Seibold F, Seibold-Schmid B, Cong Y, et al. Regional differences in L-selectin expression in murine intestinal lymphocytes. Gastroenterology. 1998; 114(5):965-974. (Biology). View Reference
-
Shortman K, Wilson A, Van Ewijk W, Scollay R. Phenotype and localization of thymocytes expressing the homing receptor-associated antigen MEL-14: arguments for the view that most mature thymocytes are located in the medulla. J Immunol. 1987; 138(2):342-351. (Biology). View Reference
-
Siegelman MH, Cheng IC, Weissman IL, Wakeland EK. The mouse lymph node homing receptor is identical with the lymphocyte cell surface marker Ly-22: role of the EGF domain in endothelial binding. Cell. 1990; 61(4):611-622. (Biology). View Reference
-
Sprent J, Tough DF. Lymphocyte life-span and memory. Science. 1994; 265(5177):1395-1400. (Biology). View Reference
-
Vestweber D. Ligand-specificity of the selectins. J Cell Biochem. 1996; 61(4):585-591. (Biology). View Reference
-
Yang G, Mizuno MT, Hellstrom KE, Chen L. B7-negative versus B7-positive P815 tumor: differential requirements for priming of an antitumor immune response in lymph nodes. J Immunol. 1997; 158(2):851-858. (Biology). View Reference
Please refer to Support Documents for Quality Certificates
Global - Refer to manufacturer's instructions for use and related User Manuals and Technical data sheets before using this products as described
Comparisons, where applicable, are made against older BD Technology, manual methods or are general performance claims. Comparisons are not made against non-BD technologies, unless otherwise noted.
For Research Use Only. Not for use in diagnostic or therapeutic procedures.