
Preparation And Storage
Recommended Assay Procedures
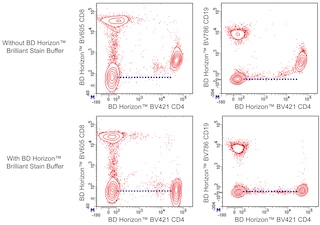
For optimal and reproducible results, BD Horizon Brilliant Stain Buffer should be used anytime two or more BD Horizon Brilliant dyes (including BD OptiBuild Brilliant reagents) are used in the same experiment. Fluorescent dye interactions may cause staining artifacts which may affect data interpretation. The BD Horizon Brilliant Stain Buffer was designed to minimize these interactions. More information can be found in the Technical Data Sheet of the BD Horizon Brilliant Stain Buffer (Cat. No. 563794).
Product Notices
- This antibody was developed for use in flow cytometry.
- The production process underwent stringent testing and validation to assure that it generates a high-quality conjugate with consistent performance and specific binding activity. However, verification testing has not been performed on all conjugate lots.
- Researchers should determine the optimal concentration of this reagent for their individual applications.
- An isotype control should be used at the same concentration as the antibody of interest.
- Caution: Sodium azide yields highly toxic hydrazoic acid under acidic conditions. Dilute azide compounds in running water before discarding to avoid accumulation of potentially explosive deposits in plumbing.
- For fluorochrome spectra and suitable instrument settings, please refer to our Multicolor Flow Cytometry web page at www.bdbiosciences.com/colors.
- Please refer to www.bdbiosciences.com/us/s/resources for technical protocols.
- BD Horizon Brilliant Stain Buffer is covered by one or more of the following US patents: 8,110,673; 8,158,444; 8,575,303; 8,354,239.
- BD Horizon Brilliant Ultraviolet 661 is covered by one or more of the following US patents: 8,110,673; 8,158,444; 8,227,187; 8,575,303; 8,354,239.
Companion Products



The 145-2C11 monoclonal antibody specifically binds to the 25-kDa ε chain of the T-cell receptor-associated CD3 complex that is expressed on thymocytes, mature T lymphocytes, and NK-T cells. The cytoplasmic domain of CD3e participates in the signal transduction events that activate several cellular biochemical pathways as a result of antigen recognition. Soluble 145-2C11 antibody can activate either unprimed (naive) or primed (memory/preactivated) T cells in vivo or in vitro, in the presence of Fc receptor-bearing accessory cells. In contrast, plate-bound 145-2C11 can activate T cells in the absence of accessory cells. Soluble 145-2C11 antibody has been reported to induce re-directed lysis of Fc receptor-bearing target cells by CTL clones and can also block lysis of specific target cells by antigen-specific CTL's. Under some conditions, T-cell activation by 145-2C11 antibody has been reported to result in apoptotic cell death. The 145-2C11 antibody does not cross-react with rat leukocytes. Preincubation of thymus cell suspensions at 37°C for 2-4 hours prior to staining reportedly enhances the ability of anti-CD3ε and anti-αβ TCR mAbs to detect the T-cell receptor on immature thymocytes.
The antibody was conjugated to BD Horizon™ BUV661 which is part of the BD Horizon Brilliant™ Ultraviolet family of dyes. This dye is a tandem fluorochrome of BD Horizon BUV395 with an Ex Max of 348-nm and an acceptor dye with an Em Max at 661-nm. BD Horizon Brilliant BUV661 can be excited by the ultraviolet laser (355 nm) and detected with a 670/25 filter and a 630 nm LP. Due to cross laser excitation of this dye, there may be significant spillover into channels detecting APC-like emissions (eg, 670/25-nm filter).
Due to spectral differences between labeled cells and beads, using BD™ CompBeads can result in incorrect spillover values when used with BD Horizon BUV661 reagents. Therefore, the use of BD CompBeads or BD CompBeads Plus to determine spillover values for these reagents is not recommended. Different BUV661 reagents (eg, CD4 vs. CD45) can have slightly different fluorescence spillover therefore, it may also be necessary to use clone-specific compensation controls when using these reagents.
Development References (14)
-
Castro JE, Listman JA, Jacobson BA, et al. Fas modulation of apoptosis during negative selection of thymocytes. Immunity. 1996; 5(6):617-627. (Clone-specific: Activation, Apoptosis). View Reference
-
Duke RC, Cohen JJ, Boehme SA, et al. Morphological, biochemical, and flow cytometric assays of apoptosis. In: Coligan J, Kruisbeek AM, Margulies D, Shevach EM, Strober W, ed. Current Protocols in Immunology. New York: John Wiley and Sons; 1995:3.17.1-3.17.33.
-
Ernst DN, Weigle WO, McQuitty DN, Rothermel AL, Hobbs MV. Stimulation of murine T cell subsets with anti-CD3 antibody. Age-related defects in the expression of early activation molecules. J Immunol. 1989; 142(5):1413-1421. (Clone-specific: Activation, Functional assay, Stimulation). View Reference
-
Isakov N, Wange RL, Burgess WH, Watts JD, Aebersold R, Samelson LE. ZAP-70 binding specificity to T cell receptor tyrosine-based activation motifs: the tandem SH2 domains of ZAP-70 bind distinct tyrosine-based activation motifs with varying affinity. J Exp Med. 1995; 181(1):375-380. (Biology). View Reference
-
Kruisbeek AM, Shevach EM. Proliferative assays for T cell function. Curr Protoc Immunol. 2004; 3:3.12.1-3.12.14. (Methodology: Activation, Stimulation). View Reference
-
Kubo RT, Born W, Kappler JW, Marrack P, Pigeon M. Characterization of a monoclonal antibody which detects all murine alpha beta T cell receptors. J Immunol. 1989; 142(8):2736-2742. (Clone-specific: Activation, Flow cytometry, Immunoprecipitation, Stimulation). View Reference
-
Leo O, Foo M, Sachs DH, Samelson LE, Bluestone JA. Identification of a monoclonal antibody specific for a murine T3 polypeptide. Proc Natl Acad Sci U S A. 1987; 84(5):1374-1378. (Immunogen: Activation, Blocking, Cytotoxicity, Flow cytometry, Immunoprecipitation, Inhibition, Stimulation). View Reference
-
Nakano H, Yamazaki T, Miyatake S, Nozaki N, Kikuchi A, Saito T. Specific interaction of topoisomerase II beta and the CD3 epsilon chain of the T cell receptor complex. J Biol Chem. 1996; 271(11):6483-6489. (Clone-specific: Functional assay, Stimulation). View Reference
-
Portoles P, Rojo J, Golby A, et al . Monoclonal antibodies to murine CD3 epsilon define distinct epitopes, one of which may interact with CD4 during T cell activation. J Immunol. 1989; 142(12):4169-4175. (Clone-specific: Blocking, Cytotoxicity, Immunoprecipitation, Radioimmunoassay). View Reference
-
Radvanyi LG, Mills GB, Miller RG. Religation of the T cell receptor after primary activation of mature T cells inhibits proliferation and induces apoptotic cell death. J Immunol. 1993; 150(12):5704-5715. (Clone-specific: Activation, Apoptosis). View Reference
-
Salvadori S, Gansbacher B, Pizzimenti AM, Zier KS. Abnormal signal transduction by T cells of mice with parental tumors is not seen in mice bearing IL-2-secreting tumors. J Immunol. 1994; 153(11):5176-5182. (Clone-specific: Activation, Calcium Flux, Flow cytometry, Western blot). View Reference
-
Shinkai Y, Alt FW. CD3 epsilon-mediated signals rescue the development of CD4+CD8+ thymocytes in RAG-2-/- mice in the absence of TCR beta chain expression. Int Immunol. 1994; 6(7):995-1001. (Biology). View Reference
-
Ucker DS, Meyers J, Obermiller PS. Activation-driven T cell death. II. Quantitative differences alone distinguish stimuli triggering nontransformed T cell proliferation or death. J Immunol. 1992; 149(5):1583-1592. (Clone-specific: Activation, Apoptosis). View Reference
-
Wang R, Murphy KM, Loh DY, Weaver C, Russell JH. Differential activation of antigen-stimulated suicide and cytokine production pathways in CD4+ T cells is regulated by the antigen-presenting cell. J Immunol. 1993; 150(9):3832-3842. (Clone-specific: Activation, Apoptosis). View Reference
Please refer to Support Documents for Quality Certificates
Global - Refer to manufacturer's instructions for use and related User Manuals and Technical data sheets before using this products as described
Comparisons, where applicable, are made against older BD Technology, manual methods or are general performance claims. Comparisons are not made against non-BD technologies, unless otherwise noted.
For Research Use Only. Not for use in diagnostic or therapeutic procedures.