
Preparation And Storage
Product Notices
- Since applications vary, each investigator should titrate the reagent to obtain optimal results.
- An isotype control should be used at the same concentration as the antibody of interest.
- Warning: Some APC-Cy7 and PE-Cy7 conjugates show changes in their emission spectrum with prolonged exposure to formaldehyde. If you are unable to analyze fixed samples within four hours, we recommend that you use BD™ Stabilizing Fixative (Cat. No. 338036).
- Cy is a trademark of Amersham Biosciences Limited. This conjugated product is sold under license to the following patents: US Patent Nos. 5,486,616; 5,569,587; 5,569,766; 5,627,027.
- This product is subject to proprietary rights of Amersham Biosciences Corp. and Carnegie Mellon University and made and sold under license from Amersham Biosciences Corp. This product is licensed for sale only for research. It is not licensed for any other use. If you require a commercial license to use this product and do not have one return this material, unopened to BD Biosciences, 10975 Torreyana Rd, San Diego, CA 92121 and any money paid for the material will be refunded.
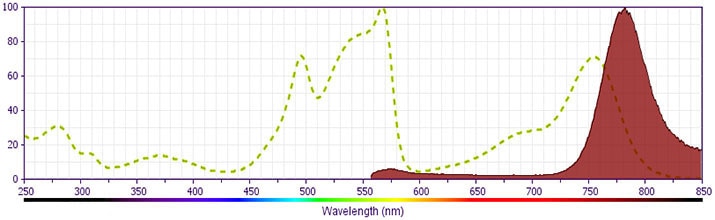
- PE-Cy7 is a tandem fluorochrome composed of R-phycoerythrin (PE), which is excited by 488-nm light and serves as an energy donor, coupled to the cyanine dye Cy7, which acts as an energy acceptor and fluoresces maximally at 780 nm. PE-Cy7 tandem fluorochrome emission is collected in a detector for fluorescence wavelengths of 750 nm and higher. Although every effort is made to minimize the lot-to-lot variation in the efficiency of the fluorochrome energy transfer, differences in the residual emission from PE may be observed. Therefore, we recommend that individual compensation controls be performed for every PE-Cy7 conjugate. PE-Cy7 is optimized for use with a single argon ion laser emitting 488-nm light, and there is no significant overlap between PE-Cy7 and FITC emission spectra. When using dual-laser cytometers, which may directly excite both PE and Cy7, we recommend the use of cross-beam compensation during data acquisition or software compensation during data analysis.
- Caution: Sodium azide yields highly toxic hydrazoic acid under acidic conditions. Dilute azide compounds in running water before discarding to avoid accumulation of potentially explosive deposits in plumbing.
- Please observe the following precautions: Absorption of visible light can significantly alter the energy transfer occurring in any tandem fluorochrome conjugate; therefore, we recommend that special precautions be taken (such as wrapping vials, tubes, or racks in aluminum foil) to prevent exposure of conjugated reagents, including cells stained with those reagents, to room illumination.
- For fluorochrome spectra and suitable instrument settings, please refer to our Multicolor Flow Cytometry web page at www.bdbiosciences.com/colors.
- Please refer to www.bdbiosciences.com/us/s/resources for technical protocols.
.png?imwidth=320)
The IM7 antibody specifically recognizes an epitope on both alloantigens and all isoforms of the CD44 glycoprotein (Pgp-1, Ly-24). The standard form of CD44, lacking variable exons and referred to as CD44H or CD44s, is widely expressed on hematopoietic and non-hematopoietic cells. CD44 isoforms encoded by variable exons are expressed on epithelial cells, but only at low levels on most leukocytes. Mice with the Ly-24.1 alloantigen (e.g., BALB/c, CBA/J, DBA/1, DBA/2) have relatively large subsets of CD44H+ T lymphocytes, while Ly-24.2 strains (e.g., A, AKR, CBA/N, C3H/He, C57BL, C57BR, C57L, C58, NZB, SJL, SWR, 129) have fewer CD44H+ T cells. CD44 is a cell adhesion receptor, and its principal ligand, hyaluronate, is a common component of extracellular matrices. Differential glycosylation of CD44 influences its binding to hyaluronate. Additional ligands include the cell surface form of CD74 and the cytokine osteopontin (Eta-1). Bone marrow- and thymus-derived progenitor cells capable of repopulating the thymus express CD44. In the periphery, the level of CD44 expression increases upon activation of B lymphocytes, CD4+ T cells, and CD8+ T cells; memory cells can be recognized by their CD44[hi] phenotype. The IM7 mAb inhibits established collagen-induced arthritis in DBA/1 mice. Moreover, it prevents CNS inflammation and clinical symptoms of experimental autoimmune encephalomyelitis. In contrast, the same antibody exacerbates experimental autoimmune thyroiditis in CBA/J mice. The IM7 mAb recognizes a different epitope from that recognized by mAb KM114, and the antibody pair can be used in ELISA to detect soluble CD44. It has been observed that IM7 antibody crossreacts with human, dog, cat, horse, cow, and pig leukocytes. Anti-human CD44, clone G44-26, and IM7 antibody compete for binding to human peripheral blood lymphocytes.
Development References (26)
-
Bendelac A. Mouse NK1+ T cells. Curr Opin Immunol. 1995; 7(3):367-374. (Biology). View Reference
-
Brocke S, Piercy C, Steinman L, Weissman IL, Veromaa T. Antibodies to CD44 and integrin alpha4, but not L-selectin, prevent central nervous system inflammation and experimental encephalomyelitis by blocking secondary leukocyte recruitment. Proc Natl Acad Sci U S A. 1999; 96(12):6896-6901. (Biology). View Reference
-
Budd RC, Cerottini JC, Horvath C, et al. Distinction of virgin and memory T lymphocytes. Stable acquisition of the Pgp-1 glycoprotein concomitant with antigenic stimulation. J Immunol. 1987; 138(10):3120-3129. (Biology). View Reference
-
Camp RL, Scheynius A, Johansson C, Pure E. CD44 is necessary for optimal contact allergic responses but is not required for normal leukocyte extravasation. J Exp Med. 1993; 178(2):497-507. (Biology). View Reference
-
Godfrey DI, Kennedy J, Suda T, Zlotnik A. A developmental pathway involving four phenotypically and functionally distinct subsets of CD3-CD4-CD8- triple-negative adult mouse thymocytes defined by CD44 and CD25 expression. J Immunol. 1993; 150(10):4244-4252. (Biology). View Reference
-
Hathcock KS, Hirano H, Murakami S, Hodes RJ. CD44 expression on activated B cells. Differential capacity for CD44-dependent binding to hyaluronic acid. J Immunol. 1993; 151(12):6712-6722. (Biology). View Reference
-
Hyman R, Lesley J, Schulte R, Trotter J. Progenitor cells in the thymus: most thymus-homing progenitor cells in the adult mouse thymus bear Pgp-1 glycoprotein but not interleukin-2 receptor on their cell surface. Cell Immunol. 1986; 101(2):320-327. (Biology). View Reference
-
Ikawa T, Kawamoto H, Fujimoto S, Katsura Y. Commitment of common T/natural killer (NK) progenitors to unipotent T and NK progenitors in the murine fetal thymus revealed by a single progenitor assay. J Exp Med. 1999; 190(11):1617-1625. (Biology). View Reference
-
Katoh S, McCarthy JB, Kincade PW. Characterization of soluble CD44 in the circulation of mice. Levels are affected by immune activity and tumor growth. J Immunol. 1994; 153(8):3440-3449. (Biology). View Reference
-
Katoh S, Zheng Z, Oritani K, Shimozato T, Kincade PW. Glycosylation of CD44 negatively regulates its recognition of hyaluronan. J Exp Med. 1995; 182(2):419-429. (Biology). View Reference
-
Lesley J, Hyman R, Kincade PW. CD44 and its interaction with extracellular matrix. Adv Immunol. 1993; 54:271-335. (Biology). View Reference
-
Lesley J, Trowbridge IS. Genetic characterization of a polymorphic murine cell-surface glycoprotein. Immunogenetics. 1982; 15(3):313-320. (Biology). View Reference
-
Lynch F, Ceredig R. Mouse strain variation in Ly-24 (Pgp-1) expression by peripheral T cells and thymocytes: implications for T cell differentiation. Eur J Immunol. 1989; 19(2):223-229. (Biology). View Reference
-
MacDonald HR, Budd RC, Cerottini JC. Pgp-1 (Ly 24) as a marker of murine memory T lymphocytes. Curr Top Microbiol Immunol. 1990; 159:97-109. (Biology). View Reference
-
Matsumoto G, Nghiem MP, Nozaki N, Schmits R, Penninger JM. Cooperation between CD44 and LFA-1/CD11a adhesion receptors in lymphokine-activated killer cell cytotoxicity. J Immunol. 1998; 160(12):5781-5789. (Biology). View Reference
-
Miyake K, Medina KL, Hayashi S, Ono S, Hamaoka T, Kincade PW. Monoclonal antibodies to Pgp-1/CD44 block lympho-hemopoiesis in long-term bone marrow cultures. J Exp Med. 1990; 171(2):477-488. (Biology). View Reference
-
Murakami S, Miyake K, June CH, Kincade PW, Hodes RJ. IL-5 induces a Pgp-1 (CD44) bright B cell subpopulation that is highly enriched in proliferative and Ig secretory activity and binds to hyaluronate. J Immunol. 1990; 145(11):3618-3627. (Biology). View Reference
-
Naor D, Sionov RV, Ish-Shalom D. CD44: structure, function, and association with the malignant process. Adv Cancer Res. 1997; 71:241-319. (Biology). View Reference
-
Naujokas MF, Morin M, Anderson MS, Peterson M, Miller J. The chondroitin sulfate form of invariant chain can enhance stimulation of T cell responses through interaction with CD44. Cell. 1993; 74(2):257-268. (Biology). View Reference
-
Nedvetzki S, Walmsley M, Alpert E, Williams RO, Feldmann M, Naor D. CD44 involvement in experimental collagen-induced arthritis (CIA). J Autoimmun. 1999; 13(1):39-47. (Biology). View Reference
-
Parish NM, Brennan FR, Cooke A. Anti-CD44 treatment does not prevent the extravasation of autopathogenic T cells to the thyroid in experimental autoimmune thyroiditis. Immunology. 1999; 97(3):533-539. (Biology). View Reference
-
Sprent J, Tough DF. Lymphocyte life-span and memory. Science. 1994; 265(5177):1395-1400. (Biology). View Reference
-
Trowbridge IS, Lesley J, Schulte R, Hyman R, Trotter J. Biochemical characterization and cellular distribution of a polymorphic, murine cell-surface glycoprotein expressed on lymphoid tissues. Immunogenetics. 1982; 15:299-312. (Immunogen). View Reference
-
Weber GF, Ashkar S, Glimcher MJ, Cantor H. Receptor-ligand interaction between CD44 and osteopontin (Eta-1). Science. 1996; 271(5248):509-512. (Biology). View Reference
-
Weiss JM, Sleeman J, Renkl AC, et al. An essential role for CD44 variant isoforms in epidermal Langerhans cell and blood dendritic cell function. J Cell Biol. 1997; 137(5):1137-1147. (Biology). View Reference
-
Zheng Z, Katoh S, He Q, et al. Monoclonal antibodies to CD44 and their influence on hyaluronan recognition. J Cell Biol. 1995; 130(2):485-495. (Biology). View Reference
Please refer to Support Documents for Quality Certificates
Global - Refer to manufacturer's instructions for use and related User Manuals and Technical data sheets before using this products as described
Comparisons, where applicable, are made against older BD Technology, manual methods or are general performance claims. Comparisons are not made against non-BD technologies, unless otherwise noted.
For Research Use Only. Not for use in diagnostic or therapeutic procedures.