The 1D3 antibody reacts with CD19, a B lymphocyte-lineage differentiation antigen. CD19, a 95-kDa transmembrance glycoprotein, is a member of the immunoglobulin superfamily and is expressed throughout B-lymphocyte development from the pro-B cell through the mature B-cell stages. Terminally differentiated plasma cells do not express CD19. On the surface of mature B cells, the CD19 molecule associates with CD21 (CR-2) and CD81 (TAPA-1), and this multimolecular complex synergizes with surface immunoglobulin to promote cellular activation. Studies with CD19-deficient mice have suggested that the level of CD19 expression affects the generation and maturation of B cells in the bone marrow and periphery. B-1 lineage B cells, also known as CD5+ B cells, are drastically reduced or absent in CD19-deficient mice. Increased levels of CD19 expression correlate with increased frequencies of peritonal and splenic B-1 cells and reduced numbers of conventional B lymphocytes in the periphery. CD19 participates in B-lymphocyte development, B-cell activation, maturation of memory B cells and regulation of tolerance. CD19 has also been detected on peritoneal mast cells, co-localized with CD21/CD35, and it is proposed to play a role in complement-mediated mast-cell activation.
The antibody was conjugated to BD Horizon BUV661 which is part of the BD Horizon Brilliant™ Ultraviolet family of dyes. This dye is a tandem fluorochrome of BD Horizon BUV395 with an Ex Max of 348-nm and an acceptor dye with an Em Max at 661-nm. BD Horizon Brilliant BUV661 can be excited by the ultraviolet laser (355 nm) and detected with a 670/25 filter and a 630 nm LP. Due to cross laser excitation of this dye, there may be significant spillover into channels detecting APC-like emissions (eg, 670/30-nm filter).
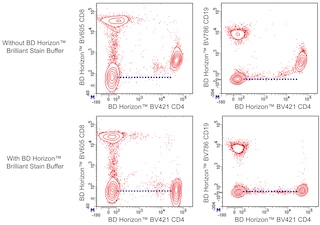
For optimal and reproducible results, BD Horizon Brilliant Stain Buffer should be used anytime two or more BD Horizon Brilliant dyes are used in the same experiment. Fluorescent dye interactions may cause staining artifacts which may affect data interpretation. The BD Horizon Brilliant Stain Buffer was designed to minimize these interactions. More information can be found in the Technical Data Sheet of the BD Horizon Brilliant Stain Buffer (Cat. No. 563794).
Due to spectral differences between labeled cells and beads, using BD™ CompBeads can result in incorrect spillover values when used with BD Horizon BUV661 reagents. Therefore, the use of BD CompBeads or BD CompBeads Plus to determine spillover values for these reagents is not recommended. Different BUV661 reagents (eg, CD4 vs. CD45) can have slightly different fluorescence spillover therefore, it may also be necessary to use clone-specific compensation controls when using these reagents.