
Dendritic cells (DCs) constitute a diverse set of hematopoietic cell types that act as conduits between innate and adaptive immune systems. They arise from lympho-myeloid hematopoiesis and are derived from the bone marrow. They are innate immune cells as they can recognize pathogens, but they can also prepare and present antigens in the context of major histocompatibility complex (MHC) proteins to prime naïve T cells to respond to threats. At least three types of DCs have been recognized: plasmacytoid DC (pDC) and myeloid/conventional DC (cDC).1 They play a key role in the tumor microenvironment.2 There is great interest in exploiting DCs to develop immunotherapies for cancer, chronic infections and autoimmune disease as well as for induction of transplantation tolerance. BD continues to expand its instrument and reagent portfolio to enable the enrichment, sorting and analysis of DCs and their different subsets by multicolor flow cytometry.
Biology of dendritic cells
DCs constitute a diverse set of hematopoietic cell types that play important roles in innate and adaptive immunity.3-5 They are potent antigen sensing and antigen presenting cells (professional APCs) that are uniquely capable of initiating primary immune responses to foreign antigens while safeguarding tolerance to self antigens.6 DCs guide the specificity, magnitude and polarity of immune responses.
Dendritic cell maturation
Immature DCs arise from progenitor cells in the bone marrow and migrate to practically all lymphoid and nonlymphoid tissues throughout the body, including the skin, lungs and intestines.7,8 A diverse array of transcription factors, signaling molecules, growth factors, cytokines, chemokines and adhesion receptors has been implicated in the differentiation pathway from common DC progenitors to mature DCs.3,9,10 In addition, through diverse assortments of surface pattern recognition receptors (PRRs), including Toll-like receptors (TLRs) and C-type lectin receptors (CLRs), immature DCs receive and process further maturation signals by discerning damage-associated molecular patterns (DAMPs) or pathogen-associated molecular patterns (PAMPs) in their local environments.3,11 This sensing of damaged cells or pathogens allows DCs to carry out their sentinel-like functions to maintain the body’s integrity.
Maturing tissue DCs alter their surface chemokine receptor and adhesion molecule profiles according to microenvironmental cues and home into secondary lymphoid organs in response to chemotactic signals. Within lymphoid tissues, immature resident or incoming nonresident DCs can be further stimulated and differentiate to become mature, functional DCs. Mature DCs have advanced capabilities to process and present antigens in the context of self-MHC antigens to naïve CD4 + or CD8 + T cells. This leads to either initiation of primary immune responses against foreign antigens or downregulation of potential T cell reactivity directed against self antigens. Mature DCs stimulate naïve T cells through their increased surface expression of peptide-loaded major histocompatibility complex (MHC) antigens, costimulatory (or coinhibitory) receptors and ligands, for example, CD80 and CD86, and the release of cytokines such as IL-6, IL-12p70 or interferons (IFNs).11,12 T cells can further tune the nature of mature DCs. Responding T cells may reciprocally regulate DCs, for example, through CD40-CD40L interactions or by T cell–derived cytokines such as IL-4 or IFN-γ. In this way, T cells may additionally instruct the professional APCs, which can promote different types of T cell–dependent immunity or tolerance.

Multifunctional roles of dendritic cells
Not only are DCs potent initiators of immune responses, they also play important regulatory roles in determining the type, magnitude and duration of immune responses that ensue.1,4,10,13 DCs accomplish this by their differential expression of cell surface ligands and receptors as well as by secreting distinct profiles of cytokines, chemokines and inflammatory mediators. For example, DCs that release IL-12p70 may preferentially promote type-1 CD4 + helper T cells (Th1) or cytolytic CD8 + T cells. Other DC types may promote T cell–dependent humoral or cell-mediated immune responses characteristic of Th2, Th9, Th17, Th22, T follicular helper (Tfh) or regulatory T (Treg) cells. The issue of exactly which DCs orchestrate these types of T cell–dependent immune responses, and how they do it, remains open and intensively investigated.
Some studies point to the DC’s maturity level as crucial, whereas others point to the major influence of the pathogen type or the tissue site involved. These are all critical parameters that require careful study. The truth may lie somewhere in between since there is such a large degree of functional plasticity within the DC pathway.14,15,16 The essential link that DCs provide between innate and adaptive immunity is also becoming more appreciated. Not only do DCs mature in response to danger signals, thus becoming capable of inducing a productive T cell response, they also trigger natural responses to invading infectious agents by activating macrophages, natural killer (NK) cells, natural killer T cells (NKT cells), granulocytes and mast cells.14 The discovery that plasmacytoid DCs (pDCs) are a major source of IFNs, quickly secreting them in response to certain viruses,9 serves as an important example of the multifunctional role played by DCs in both innate and adaptive immune responses.
Dendritic cell heterogeneity
Multiple types of precursor, immature and mature DCs (for example, Langerhans cells, dermal or interstitial DCs, blood DCs) that differ in origin, morphology, localization, maturation state, phenotype and function10,14 have been described. Despite some cell surface phenotypic differences between the two species, two generally accepted types of DCs have been described in human and mouse model systems that appear to represent different lineages: plasmacytoid DCs (pDCs) and myeloid DCs (mDCs), also known as classical or conventional DCs (cDCs).1 pDCs have a tremendous capacity to produce IFNs but may not present antigens as efficiently as mDCs.1,3 Human pDCs are distinguished by their coexpression of CD123 and CD304 whereas mouse pDCs express CD45R/B220 and Ly-6C.1,9,10 Two major classes of mDCs have been further classified in the human and mouse species, which are defined by the alternative expression of either IFN regulatory factor 4 (IRF4+ DCs) or IRF-8 (IRF-8+ DCs).13 IRF4+ DCs in humans characteristically express CD1c, whereas mouse counterparts express either CD4 (lymphoid resident DCs) or CD11b (migratory DCs). The IRF4+ DCs from both species coexpress CD172a/Sirp-α and can efficiently present antigens to naïve CD4+ T cells. Conversely, human IRF8+ DCs typically express CD141, while mouse equivalents express CD8a (lymphoid resident DCs) or CD103 (migratory DCs) with all subsets expressing the XCR1 chemokine receptor, CD370/Clec9a, and capable of presenting antigen to CD4+ T cells and CD8+ T cells. Human and mouse Langerhans cells (LCs) likewise coexpress several distinguishing markers in common including CD207/Langerin, CD326/EpCAM and CD324/E-Cadherin.3,10 DC subsets residing in the dermis and intestines of both species have also been described.3 For a summary of human and mouse DC counterparts, see the table below.
Functionality of human DC subsets and their mouse DC counterparts
| Human DC Subsets | Mouse DC Counterparts | Frequency | Localization | Cytokine Production Upon Stimulation* |
|---|---|---|---|---|
| pDC | pDC | ~1% peripheral blood mononuclear cells (PBMCs) | Human blood Lymph node Tcell zone Tonsil |
IFN-I+, IFN-III (IFN-λ)+ IL-6+, IL-8+ IP-10 (CXCL10)+ TNF+ |
| CD1c+ DCs | CD4+ or CD11b+ DCs | ~1% PBMCs | Human blood Nonlymphoid tissues: Skin, liver, lung, and gut Lymphoid tissues: spleen, lymph nodes |
IL-1β+, IL-6+, IL-8+ IL-10+, IL-12+ IL-23+ TNF+ IL-15+ (skin) |
| CD141+ | CD8+ or CD103+ DCs | 0.03% PBMCs CD8+ DCs: 20–40% of mouse spleen and lymph node cDCs | Human lymph node, tonsil, spleen, bone marrow Human nonlymphoid tissues: skin, lung, liver, intestine CD8+ DCs: Mouse lymphoid tissues |
IFN-I+, IFN-III (IFN-λ)+ IL-12+ (mouse) CXCL-10 (IP-10)+ TNF+** |
| LCs (Langerhans cells) | LCs (Langerhans cells) | 3–5% epidermal cells | Human stratified squamous epithelia, draining lymph nodes | IL-15+ |
| Inflammatory DCs | Inflammatory DCs | Inflammatory sites | IL-1β+, IL-6+ IL-10+, IL-12+, IL-23+ TNF+ |
*Cytokine production could vary with the stimulant used, the stimulation conditions or the physiological state of the cell.
**TNF is not typically produced by human CD141+ DCs in response to TLR8 stimulation.
Another class of DCs, inflammatory DCs, may arise from monocytes that may be driven by environmental stimuli to take on the characteristics and functions of DCs.3 Clearly, provocative interspecies differences as well as similarities in certain functionally related molecules are being described for the various DC subsets including their expressed profiles of TLRs, CLRs, CD1 molecules, chemokine receptors and their cytokine secretion patterns.3 Since a combination of factors, including the DC subset and maturation stage, influence resulting T cell responses, detailed phenotypic analysis combined with functional studies will be one of the useful approaches in further studying the intricacies of DC biology in physiological as well as pathological conditions.
References
- Palucka K and Banchereau J. Cancer immunotherapy via dendritic cells. Nat Rev Cancer. 2012;12(4):265-277. doi: 10.1038/nrc3258
- Janco JMT, Lamichhane P, Karyampudi L, Knutson K. Tumor-infiltrating dendritic cells in cancer pathogenesis. J Immunol. 2015;194(7):2985-2991. doi: 10.4049/jimmunol.1403134
- Merad M, Sathe P, Helft J, Miller J, Mortha A. The dendritic cell lineage: ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Annu Rev Immunol. 2013;31:563-604. doi: 10.1146/annurev-immunol-020711-074950
- Schraml BU, Reis e Sousa C. Defining dendritic cells. Curr Opin Immunol. 2015;32:13-20. doi: 10.1016/j.coi.2014.11.001
- Collin M, McGovern N, Haniffa M. Human dendritic cell subsets. Immunology. 2013;140(1):22-30. doi: 10.1111/imm.12117
- O'Keeffe M, Mok WH, Radford KJ. Human dendritic cell subsets and function in health and disease. Cell Mol Life Sci. 2015;72(22):4309-4325. doi: 10.1007/s00018-015-2005-0
- Apostolopoulos V, Thalhammer T, Tzakos AG, Stojanovska L. Targeting antigens to dendritic cell receptors for vaccine development. J Drug Deliv. 2013;2013:869718. doi: 10.1155/2013/869718
- Cohn L, Delamarre L. Dendritic cell-targeted vaccines. Front Immunol. 2014;5:255. doi: 10.3389/fimmu.2014.00255
- Delamarre L, Mellman I. Harnessing dendritic cells for immunotherapy. Sem Immunol. 2011;23(1):2-11. doi: 10.1016/j.smim.2011.02.001
- Breton G, Lee J, Liu K, Nussenzweig MC. Defining human dendritic cell progenitors by multiparametric flow cytometry. Nat Protoc.2015;10(9):1407-1422. doi: 10.1038/nprot.2015.092
- Swiecki M, Colonna M. The multifaceted biology of plasmacytoid dendritic cells. Nat Rev Immunol. 2015;15(8):471-485. doi: 10.1038/nri3865
- Murphy TL, Grajales-Reyes GE, Wu X, et al. Transcriptional control of dendritic cell development. Annu Rev Immunol. 2016;34:93-119. doi: 10.1146/annurev-immunol-032713-120204
- Poltorak MP, Schraml BU. Fate mapping of dendritic cells. Front Immunol. 2015;6:199. doi: 10.3389/fimmu.2015.00199
- Dutertre CA, Wang LF, Ginhoux F. Aligning bona fide dendritic cell populations across species. Cell Immunol. 2014;291(1-2):3-10. doi: 10.1016/j.cellimm.2014.08.006
- Schlitzer A, Ginhoux F. Organization of the mouse and human DC network. Curr Opin Immunol. 2014;26:90-99. doi: 10.1016/j.coi.2013.11.002
- Reis e Sousa C. Activation of dendritic cells: translating innate into adaptive immunity. Curr Opin Immunol. 2004;16(1):21-25. doi: 10.1016/j.coi.2003.11.007.
Human dendritic cells
Human dendritic cell subsets can be characterized by a number of surface and intracellular markers through flow cytometry.
The following examples on peripheral blood illustrate DC subset identification based on available BD OptiBuild™ Custom Reagents.

See the table below for key markers for phenotypic characterization of various human DC subsets; intracellular markers are indicated in bold. BD Life Sciences has a broad portfolio of antibody reagents to most of these specificities in multiple formats to allow flexibility in panel design and downstream analysis.
All human DC subsets are identified as lineage– (CD3– CD19– CD20– CD56– CD141) and CD45+ MHCII (HLA-DR)+ CD11c+/-2.
| DC Subset | Key Markers | |||
|---|---|---|---|---|
| Primary Markers | Additional Positive Markers | Additional Negative Markers | Transcription Factors | |
| Plasmacytoid DCs | CD123 (IL-3Rα)high CD303 (BDCA2/CLEC4C)+ CD304 (Neuropilin-1/BDCA4)+ CD85g (ILT7)+ CD11clow CD11b–, CD14– |
CD2±, CD4+, CD45RA+ CD141 (BDCA3)low CD272 (BTLA)+ CD366 (TIM-3)± CD367 (DCIR/CLEC4A)+ CD371 (CLEC12A)± TLR7high, TLR9high FLT3 (CD135)+ GM-CSFR (CD116)+ CCR5+, CXCR4+ |
CD1a–, CD1c (BDCA1)– CD16 (FcγRIII)– CD172a (Sirp-α)– CD207 (Langerin)– CD324 (E-Cadherin)– CD326 (EpCAM)– CD369 (Dectin-1/CLEC7A)– CD370 (CLEC9A/DNGR1)– CLEC6A (Dectin-2)– XCR1– |
IRF7+, IRF8+ SpiB+ |
| CD1c+ Myeloid DCs |
CD1c (BDCA1)+ CD172a (Sirp-α)+ CLEC6A (Dectin-2)+CD11b+/low, CD11c+ CX3CR1+CD14low/– |
CD4+, CD13+, CD26low, CD33+ CD45RO+, CD141 (BDCA3)± CD272(BTLA)+ CD366 (TIM-3)+ CD367 (DCIR/CLEC4A)+ CD369 (Dectin-1/CLEC7A)+ CD371 (CLEC12A)+ CD1a+ [skin and intestine] CD141(BDCA3)+ [intestine] TLR3low, TLR4low, TLR8+, TLR10low FLT3 (CD135)+, GM-CSFR (CD116)+ |
CD1a–, CD16 (FcγRIII)– CD123 (IL-3Rα)– CD207 (Langerin)– CD304 (Neuropilin-1/BDCA4)–, CD324 (E-Cadherin)– CD326 (EpCAM)– CD370 (CLEC9A/DNGR1)– ESAM– XCR1– | IRF4+ |
| CD141+ Myeloid DCs |
CD141 (BDCA3)high CD370 (CLEC9A/DNGR1)+ NECL2 (CADM1)+ CD11c+/low CD14– |
CD4+, CD11blow CD26+, CD162high CD205 (DEC-205)high CD272 (BTLA)high CD367 (DCIR/CLEC4A)+ CD366 (TIM-3)+ CD369 (Dectin-1/CLEC7A)+ CD371 (CLEC12A)+ TLR3+, TLR8+ FLT3 (CD135)+ GM-CSFR (CD116)+ XCR1+ |
CD1a−, CD1c (BDCA1)−, CD16 (FcγRIII)− CD172a (Sirp–α)− CD207 (Langerin)− CD304 (Neuropilin-1/BDCA4)− CD324 (E-Cadherin)− CD326 (EpCAM)− |
IRF8+ BATF3+ |
| Langerhans Cells | CD207 (Langerin)+ CD324 (E-Cadherin)+ CD326 (EpCAM)+ CD11blow, CD11c+ CD14− |
CD1ahigh, CD1c (BDCA1)+ CD36+ CD172a (Sirp-α)+ CD369 (Dectin-1/CLEC7A)+, CD371 (CLEC12A)+ CLEC6A (Dectin-2)+ TLR1+, TLR2+, TLR3low, TLR6+ |
CD304 (Neuropilin-1/BDCA4)– CD367 (DCIR/CLEC4A)– XCR1– |
|
| CD1a+ Dermal DCs |
CD1a+ CD64 (FcγRI)+ CD366 (TIM-3)+ CD11b+, CD11c+ CD14− |
CD1c (BDCA1)+ CD172a (Sirp-α)+ CD367 (DCIR/CLEC4A)+ CD369 (Dectin-1/CLEC7A)+ CLEC6A (Dectin-2)+ TLR1–3+, TLR6+, TLR7+, TLR10+ |
CD207 (Langerin)− CD209 (DC-SIGN/CLEC4L)− CD324 (E-Cadherin)− CD326 (EpCAM)− |
|
| CD14a+ Dermal DCs |
CD14+ CD209 (DC-SIGN/CLEC4L)+ CD11b+, CD11c+ |
CD1c (BDCA1)+ CD172a (Sirp-α)+ CD367 (DCIR/CLEC4A)+ CD369 (Dectin-1/CLEC7A)+ CLEC6A (Dectin-2)+ TLR1–3+, TLR6, TLR7+ CSF–1R (CD115)+ |
CD1a− CD207 (Langerin)− CD324 (E-Cadherin)− CD326 (EpCAM)− CD366 (TIM-3)− |
|
| Inflammatory DCs (Monocyte-Derived DCs) |
CD16 (FcγRIII)+ CD64 (FcγRI)+ CD1a+ CD1c (BDCA1)+ CD11b+, CD11c+ CD14± |
CD172a (Sirp-α)+ CD206 (MR/CLECL13D)+ CD209 (DC-SIGN/CLEC4L)+ CD367 (DCIR/CLEC4A)+ CD369 (Dectin-1/CLEC7A)+ CD371 (CLEC12A)+ CLEC6A (Dectin-2)+ TLR3low, TLR4+, TLR7low, TLR8+ CCR2 (CD192)+ |
CD207 (Langerin)− CD324 (E-Cadherin)− CD326 (EpCAM)− |
1CD14 is negative or low on all DC subsets except for CD14+ dermal DCs and inflammatory DCs.
2CD11c is positive on all DC subsets except for plasmacytoid DCs where expression has been reported as low or negative.
TLR7, 8 and 9 are endosomal and require intracellular staining.
Mouse dendritic cells
Mouse dendritic cell (DC) subsets can be characterized by a number of surface and intracellular markers through flow cytometry.
The following examples on mouse spleen illustrate DC subset identification based on available BD OptiBuild™ Custom Reagents.
Immunophenotyping of mouse dendritic cell subsets. Ten-color immunophenotypic characterization of the three main subsets of mouse DCs (myeloid, lymphoid and plasmacytoid) in the mouse spleen. BALB/c mouse spleen was enzymatically digested, stained with a cocktail of antibodies (including BD OptiBuild™ Reagents Sirpα BV650 and Clec12A BV786) and analyzed on a BD FACSCelesta™ Flow Cytometer Blue/Violet/ Red (BVR) laser configuration.
Cells were initially gated on CD3, CD19 and 7-AAD negative cells (not shown). A-D. Gating strategy: I-A/I-EhighCD11chigh conventional dendritic cells (DCs) were further discriminated into CD11b+ mDCs and CD8+ lDCs. I-A/I-ElowCD11clow B220+ Gr1+ cells were recognized as pDCs. Results: Differential expression of the additional markers CD4, CD172a (Sirpα) and CD371 (Clec12A) was further analyzed within the mDC (E, F, G), lDC (H, I, J) and pDC (K, L, M) subsets, respectively. Gates were drawn based on fluorescence minus one (FMO) controls.
See the table below for key markers for phenotypic characterization of various mouse DC subsets; intracellular markers are indicated in bold. BD Life Sciences has a broad portfolio of antibody reagents to most of these specificities in multiple formats to allow flexibility in panel design and downstream analysis.
Mouse DC markers are identified as lineage – (CD3– CD19– CD49b– or NK1.1– CD14–) and CD45+ MHCII+ CD11c+.

| DC Subset | Key Markers | |||
|---|---|---|---|---|
| Primary Markers | Additional Positive Markers | Additional Negative Markers | Transcription Factors | |
| Plasmacytoid DCs (pDCs) |
CD45R (B220)+ CD317 (BST-2)+ Ly-6C+ Siglec-H+ CD11clow CD14− |
CD4+, CD26+ CD172a (Sirp-α)+ CD209a (DC-SIGN)+ CD272 (BTLA)+ CD370 (Clec9a/DNGR1)+ Gr1 (Ly-6C and Ly-6G)+ FLT3 (CD135)+ TLR7high, TLR9high |
CD11b− CD24− CD36−, CD64 (FcγRI)−, CD103− CD205 (DEC-205)− CD207 (Langerin)− CD326 (EpCAM)− DCIR2 (Clec4a4/33D1)− F4/80− CX3CR1−, XCR1− |
IRF7+, IRF8+ BATF3high SpiB+ Zbtb46− |
| CD4+ CD11b+ Lymphoid-Resident DCs |
CD4+ CD11b+ CD11chigh CD8a− CD14− |
CD24+, CD26+ CD172a (Sirp-α)+ CD205 (DEC-205)+ CD209a (DC-SIGN)+ CD272 (BTLA)low DCIR2 (Clec4a4/33D1)+ ESAM+ F4/80+ FLT3 (CD135)+ TLR5+, TLR7+, TLR9+ CX3CR1+ |
CD36− CD45R (B220)− CD64 (FcγRI)−, CD103− CD207 (Langerin)− CD326 (EpCAM)− CD370 (Clec9a/DNGR1)− Ly6C− XCR1− |
IRF4+ Zbtb46+ BATF3high |
| CD4− CD11b+ Conventional Migratory DCs |
CD11b+ CD11c+ CD4− CD8a− CD14− |
CD24±, CD26+ CD64 (FcγRI)+ CD172a (Sirp-α)+ CD209a (DC-SIGN)± CD272 (BTLA)low DCIR2 (Clec4a4/33D1)+ F4/80+ Ly-6C± FLT3 (CD135)+ TLR5+, TLR7+, TLR9+ CX3CR1+ |
CD36− CD45R (B220)−, CD103− CD207 (Langerin)− CD326 (EpCAM)− CD370 (Clec9a/DNGR1)− XCR1− |
IRF4+ IRF2+ Zbtb46+ BATF3high |
| CD8a+ Conventional Lymphoid-Resident DCs |
CD8a+ CD11chigh CD4− CD11b− CD14− |
CD1d1+, CD24+, CD26+ CD36+, CD103± CD205 (DEC-205)+ CD207 (Langerin)± CD272 (BTLA)high CD370 (Clec9a/DNGR1)+ NECL2 (CADM1)+ FLT3 (CD135)+ TLR3+, TLR4+, TLR11+ XCR1+, CX3CR1± |
CD45R (B220)− CD64 (FcγRI)− CD172a (Sirp-α)− CD209a (DC-SIGN)− CD326 (EpCAM)− DCIR2 (Clec4a4/33D1)− F4/80− Ly-6C− |
IRF8+ BATF3+ Zbtb46+ |
| CD103+ Conventional Migratory DCs |
CD103+ CD11chigh CD4− CD8− CD11b− CD14− |
CD1d1+, CD24+ CD26+, CD36+ CD205 (DEC-205)+ CD207 (Langerin)+ CD272 (BTLA)high CD370 (Clec9a/DNGR1)+ NECL2 (CADM1/CD317)+ FLT3 (CD135)+ TLR3+, TLR4+, TLR11+ XCR1+ |
CD45R (B220)− CD64 (FcγRI)− CD172a (Sirp-α)− CD209a (DC-SIGN)− CD326 (EpCAM)− DCIR2 (Clec4a4/33D1)− F4/80− Ly-6C− CX3CR1− |
IRF8+ BATF3+ Zbtb46+ |
| Langerhans Cells | CD207 (Langerin)+ CD324 (E-Cadherin)+ CD326 (EpCAM)+ CD11b+ CD11c+ CD14− |
CD24+ CD172a (Sirp-α)+ CD205 (DEC-205)+ F4/80+ TLR3+, TLR11+ |
CD8a−, CD26−, CD36− CD45R (B220)−, CD103− CD209a (DC-SIGN)− CD370 (Clec9a/DNGR1)− DCIR2 (Clec4a4, 33D1)− Ly-6C− CX3CR1−, XCR1− |
|
| CD207+ Dermal DCs | CD207 (Langerin)+ CD11blow CD11c+ |
CD103± | CD45R (B220)− CD172a (Sirp-α)− CD326 (EpCAM)− |
|
| CD207− Dermal DCs | CD207 (Langerin)− CD11b± CD11c+ |
CD172a (Sirp-α)+ | CD45R (B220)− CD103− CD326 (EpCAM)− |
|
| Inflammatory DCs (Monocyte–Derived DCs) |
CD64 (FcγRI)+
CD11b+ CD11c+ CD14− |
CD209a (DC-SIGN)+ CD272 (BTLA)low Ly-6C+ TLR1-6, TLR7-8+, TLR10+ |
CD8a−, CD45R (B220)−, CD103− CD172a (Sirp-α)− CD207 (Langerin)− CD326 (EpCAM)− |
Isolation of dendritic cells
Certain research approaches require the use of isolated populations of dendritic cells (DCs) for further characterization, culture or functional studies. Isolation of these very rare cell types, which make up only 0.1–1% of blood mononuclear cells, has been a limitation historically.1 Advances in flow cytometry and magnetic-bead separation technologies have made DC isolation easier. Flow cytometry–based cell sorting enables the isolation of single cells for further analysis.
Dendritic cell populations
Isolated DC populations are often used to carry out functional assays, such as co-culture with T cells, or for other downstream applications. BD Biosciences offers approaches for the successful isolation of both human and mouse DCs.
The BD IMag™ Cell Separation System is based on a simple yet highly effective direct magnet technology. Positive or negative selection of DCs, for example, is feasible with researchers selecting from a range of BD IMag™ Particle-formatted cell surface binding antibodies. Easy-to-use human and mouse DC enrichments sets, employing a negative selection of non-DC populations in a few short steps, are also available.
Flow cytometric sorting is a very effective way to isolate these extremely rare cells, since it delivers high purity and recovery of cells and is well suited for the isolation of cells defined by a multimarker phenotype. Many researchers have relied on BD FACSAria™ Cell Sorters to isolate specific DC populations.2
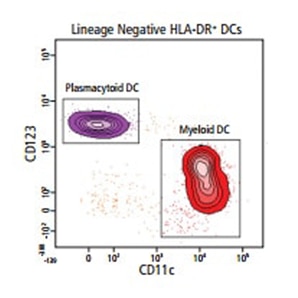
Isolation of pDCs and mDCs from human PBMCs
Plasmacytoid DCs (pDCs) and myeloid DCs (mDCs) can be isolated from human peripheral blood on the basis of the phenotypes Lin– HLA-DR+ CD123+ CD11c– (pDCs) and Lin– HLA–DR+ CD123– CD11c+(mDCs). In the example shown below, a pre-enrichment step was performed using a BD IMag™ Human Dendritic Cell Enrichment Set, Cat. No. 558420, on peripheral blood mononuclear cells (PBMCs), negatively selecting erythrocytes, platelets and peripheral leucocytes (that are not DCs), thereby providing a DC-enriched sample for cell surface staining and sorting.
Cells were stained with lineage cocktail FITC, CD123 BV421, HLA-DR PE and CD11c APC. The viability dye FVS780 was used to exclude dead cells. Cells were sorted using a BD FACSAria™ Flow Cytometer. The combination of BD IMag™ Particle-based enrichment and flow cytometric sorting is a convenient time-reducing workflow generating highly pure DC populations normally found at low frequencies in human blood.
BD Application Specialists are available to provide field- or phone-based advice to support you in your sorting applications. Visit our Support page for more information.
BD flow cytometry platforms such as the BD FACSMelody™, BD FACSAria™ III, BD FACSAria™ Fusion Cell Sorters support single-cell sorting into different types of plates. Single-cell sorting is made possible by the automated deposition unit, and this utility is enhanced when combined with the index-sorting function, which records the flow cytometric data (e.g., marker phenotype) and sort location (X and Y coordinates of the sort-collection device) for each sorted event. In this way, results of post-sorting assays, which may include DNA or RNA sequence analysis, can be precisely traced back to the flow characteristics of the specific cell.

Cytokine detection
Measurement of the types and amounts of cytokines that are secreted by dendritic cells (DCs) provides insight into the nature and magnitude of the DC response. Multiplexed assays are increasingly being used to measure secreted factors, such as cytokines and chemokines.
The broad dynamic range of fluorescence detection and multiplexed measurement allow for small sample volumes, fewer sample dilutions and substantially less time to establish the value of an unknown vs a conventional enzyme-linked immunosorbent assay (ELISA) approach.
The BD® CBA portfolio includes assays for measurement of a variety of soluble factors, including secreted proteins such as cytokines or chemokines, shed or released cell surface markers (for example, sCD14) plus cell signaling molecules such as phosphoproteins within cell lysates.
Intracellular detection of cytokines
By using a protein transport inhibitor to block secretion, cytokines can be detected in the cell in which they are being produced. Hence, it is possible to distinguish whether cytokine production by an activated cell population is the result of a few cells producing large amounts of cytokine or a large population of cells each producing small quantities of cytokine.

Transcription factors and other intracellular proteins
Analysis of DC physiology, from early development to effector functions of mature DCs, has revealed intracellular changes associated with developmental status, cellular activation or other functional properties.
BD offers solutions that enable analysis of these intracellular molecules, to support researchers in deciphering the interconnected pathways regulating DC biology.
Detection of transcription factors
BD offers several DC-relevant, flow cytometry validated transcription-factor antibodies. For example, in mice, Zbtb46 is expressed in conventional DCs and their precursors but not in pDCs. Conversely, Spi-B is required for pDC development and function.3

Detection of phosphoproteins
BD Phosflow™ Antibodies are monoclonal phosphoepitope-specific antibodies validated for flow cytometric detection. The recommended permeabilization buffer for most BD Phosflow™ Antibodies is BD Phosflow™ Perm Buffer III, but alternative permeabilization buffers are also available, for enabling cell surface staining for subpopulation analysis, for example.
A number of BD Phosflow™ Antibody specificities are available for analysis of signaling pathways involved in DC development and activation, such as reagents for pSTAT3 and pSTAT5 which have differing effects on pDC development. The BD Phosflow™ System has been used to measure phosphoproteins such as pSyk after viral infection of pDCs in vitro1 and is widely employed within cell-signaling studies for innate and adaptive immunity.

Immunofluorescence and immunohistochemistry
DC morphology and subcellular localization
Using monoclonal antibodies with fluorescence imaging allows for the identification of DCs within tissues and insights into the mechanistic aspects of DC biology in vitro.
BD monoclonal antibodies that are directly conjugated to new and bright photostable fluorescent dyes, such as BD Horizon Brilliant Violet™ 421 (BV421) and BD Horizon™ BV480, enable multicolor imaging beyond conventional three- to four-color microscopy.
")
References
- Merad M, Sathe P, Helft J, Miller J, Mortha A. The dendritic cell lineage: ontogeny and function of dendritic cells and their subsets in the steady state and the inflamed setting. Annu Rev Immunol. 2013;31:563-604. doi: 10.1146/annurev-immunol-020711-074950
- Collin M, McGovern N, Haniffa M. Human dendritic cell subsets. Immunology. 2013;140(1):22-30. doi: 10.1111/imm.12117
- Janco JMT, Lamichhane P, Karyampudi L, Knutson K. Tumor-infiltrating dendritic cells in cancer pathogenesis. J Immunol. 2015;194(7):2985-2991. doi: 10.4049/jimmunol.1403134
For Research Use Only. Not for use in diagnostic or therapeutic procedures.
Alexa Fluor is a trademark of Life Technologies Corporation.
The information provided herein is not meant to be used, nor should it be used, to diagnose or treat any medical condition. All content, including text, graphics, images and information etc., contained in or available through this literature is for general information purposes only. For diagnosis or treatment of any medical condition, please consult your physician/doctor. Becton Dickinson India Private Limited and or its affiliates, its employees are not liable for any damages/claims to any person in any manner whatsoever.