
Preparation And Storage
Recommended Assay Procedures
BD® CompBeads can be used as surrogates to assess fluorescence spillover (Compensation). When fluorochrome conjugated antibodies are bound to BD® CompBeads, they have spectral properties very similar to cells. However, for some fluorochromes there can be small differences in spectral emissions compared to cells, resulting in spillover values that differ when compared to biological controls. It is strongly recommended that when using a reagent for the first time, users compare the spillover on cells and BD CompBeads to ensure that BD® CompBeads are appropriate for your specific cellular application.
Product Notices
- Since applications vary, each investigator should titrate the reagent to obtain optimal results.
- Please refer to www.bdbiosciences.com/us/s/resources for technical protocols.
- Although hamster immunoglobulin isotypes have not been well defined, BD Biosciences Pharmingen has grouped Armenian and Syrian hamster IgG monoclonal antibodies according to their reactivity with a panel of mouse anti-hamster IgG mAbs. A table of the hamster IgG groups, Reactivity of Mouse Anti-Hamster Ig mAbs, may be viewed at http://www.bdbiosciences.com/documents/hamster_chart_11x17.pdf.
- Caution: Sodium azide yields highly toxic hydrazoic acid under acidic conditions. Dilute azide compounds in running water before discarding to avoid accumulation of potentially explosive deposits in plumbing.
- An isotype control should be used at the same concentration as the antibody of interest.
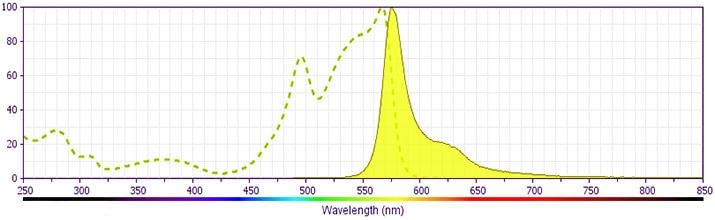
- For fluorochrome spectra and suitable instrument settings, please refer to our Multicolor Flow Cytometry web page at www.bdbiosciences.com/colors.
- Please refer to http://regdocs.bd.com to access safety data sheets (SDS).
.png?imwidth=320)
The H1.2F3 monoclonal antibody specifically binds to CD69 (Very Early Activation antigen), an 85 kDa disulfide-linked homodimer of differentially glycosylated subunits. CD69 is a C-type lectin, most closely related to the NKR-P1 and Ly-49 NK cell-activation molecules. Its expression is rapidly induced upon activation of lymphocytes (T, B, NK, and NK-T cells), neutrophils, and macrophages. CD69 is expressed also on thymocytes that are undergoing positive selection; its role in that process is unclear. H1.2F3 mAb augments PMA-induced T-cell stimulation and IFN-γ-induced macrophage stimulation. IL-2-activated NK cells express CD69, and H1.2F3 mAb induces redirected lysis of FcR-bearing target cells by NK cells.
Development References (16)
-
Bendelac A, Matzinger P, Seder RA, Paul WE, Schwartz RH. Activation events during thymic selection. J Exp Med. 1992; 175(3):731-742. (Biology). View Reference
-
Brandle D, Muller S, Muller C, Hengartner H, Pircher H. Regulation of RAG-1 and CD69 expression in the thymus during positive and negative selection. Eur J Immunol. 1994; 24(1):145-151. (Biology). View Reference
-
Gabor MJ, Godfrey DI, Scollay R. Recent thymic emigrants are distinct from most medullary thymocytes. Eur J Immunol. 1997; 27(8):2010-2050. (Biology). View Reference
-
Karlhofer FM, Yokoyama WM. Stimulation of murine natural killer (NK) cells by a monoclonal antibody specific for the NK1.1 antigen. IL-2-activated NK cells possess additional specific stimulation pathways. J Immunol. 1991; 146(10):3662-3673. (Clone-specific: Induction). View Reference
-
Keefe R, Dave V, Allman D, Wiest D, Kappes DJ. Regulation of lineage commitment distinct from positive selection. Science. 1999; 286(5442):1149-1153. (Biology). View Reference
-
Lauzurica P, Sancho D, Torres M, et al. Phenotypic and functional characteristics of hematopoietic cell lineages in CD69-deficient mice. Blood. 2000; 95(7):2312-2320. (Biology). View Reference
-
Marzio R, Jirillo E, Ransijn A, Mauel J, Corradin SB. Expression and function of the early activation antigen CD69 in murine macrophages. J Leukoc Biol. 1997; 62(3):349-355. (Clone-specific: Stimulation). View Reference
-
Merkenschlager M, Graf D, Lovatt M, Bommhardt U, Zamoyska R, Fisher AG. How many thymocytes audition for selection. J Exp Med. 1997; 186(7):1149-1158. (Biology). View Reference
-
Nishimura T, Kitamura H, Iwakabe K, et al. The interface between innate and acquired immunity: glycolipid antigen presentation by CD1d-expressing dendritic cells to NKT cells induces the differentiation of antigen-specific cytotoxic T lymphocytes. Int Immunol. 2000; 12(7):987-994. (Biology). View Reference
-
Punt JA, Suzuki H, Granger LG, Sharrow SO, Singer A. Lineage commitment in the thymus: only the most differentiated (TCRhibcl-2hi) subset of CD4+CD8+ thymocytes has selectively terminated CD4 or CD8 synthesis. J Exp Med. 1996; 184(6):2091-2099. (Biology). View Reference
-
Sobel ES, Yokoyama WM, Shevach EM, Eisenberg RA, Cohen PL. Aberrant expression of the very early activation antigen on MRL/Mp-lpr/lpr lymphocytes. J Immunol. 1993; 150(2):673-682. (Clone-specific: Stimulation). View Reference
-
Wilkinson RW, Anderson G, Owen JJ, Jenkinson EJ. Positive selection of thymocytes involves sustained interactions with the thymic microenvironment. J Immunol. 1995; 155(11):5234-5240. (Biology). View Reference
-
Yokoyama WM, Koning F, Kehn PJ, et al. Characterization of a cell surface-expressed disulfide-linked dimer involved in murine T cell activation. J Immunol. 1988; 141(2):369-376. (Immunogen: Flow cytometry, Stimulation). View Reference
-
Yokoyama WM, Maxfield SR, Shevach EM. Very early (VEA) and very late (VLA) activation antigens have distinct functions in T lymphocyte activation. Immunol Rev. 1989; 109:153-176. (Clone-specific: Flow cytometry, Stimulation). View Reference
-
Ziegler SF, Levin SD, Johnson L, et al. The mouse CD69 gene. Structure, expression, and mapping to the NK gene complex. J Immunol. 1994; 152(3):1228-1236. (Biology). View Reference
-
Ziegler SF, Ramsdell F, Alderson MR. The activation antigen CD69. Stem Cells. 1994; 12(5):456-465. (Biology). View Reference
Please refer to Support Documents for Quality Certificates
Global - Refer to manufacturer's instructions for use and related User Manuals and Technical data sheets before using this products as described
Comparisons, where applicable, are made against older BD Technology, manual methods or are general performance claims. Comparisons are not made against non-BD technologies, unless otherwise noted.
For Research Use Only. Not for use in diagnostic or therapeutic procedures.