The RM2-5 monoclonal antibody specifically binds to the immunoglobulin superfamily adhesion molecule, CD2 (LFA-2). CD2 is a type I transmembrane glycoprotein that serves as the the major receptor for CD48 in the mouse. CD2 is involved in T-cell activation, immunoregulation, and thymocyte maturation. In the mouse, CD2 is expressed on peripheral T lymphocytes, B lymphocytes, and NK cells, and a subpopulation of intraepithelial T lymphocytes. CD2 is expressed throughout mouse thymic ontogeny, except for distinct subsets of the CD4-CD8- early thymocytes. In the mouse bone marrow, CD2 is expressed on B220+ sIg+ CD43- pre-B cells, but not on CD43+ pro-B cells. The RM2-5 antibody is one of a set of five anti-mouse CD2 monoclonal antibodies that were classified into two groups according to their mutual competition in binding to cell surface CD2 and which block CD2-mediated cell-cell adhesion.
The antibody was conjugated to BD Horizon™ BUV661 which is part of the BD Horizon Brilliant™ Ultraviolet family of dyes. This dye is a tandem fluorochrome of BD Horizon BUV395 with an Ex Max of 348-nm and an acceptor dye with an Em Max at 661-nm. BD Horizon Brilliant BUV661 can be excited by the ultraviolet laser (355 nm) and detected with a 670/25 filter and a 630 nm LP. Due to cross laser excitation of this dye, there may be significant spillover into channels detecting APC-like emissions (eg, 670/25-nm filter).
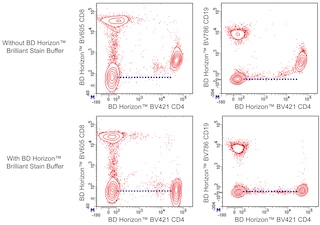
Due to spectral differences between labeled cells and beads, using BD™ CompBeads can result in incorrect spillover values when used with BD Horizon BUV661 reagents. Therefore, the use of BD CompBeads or BD CompBeads Plus to determine spillover values for these reagents is not recommended. Different BUV661 reagents (eg, CD4 vs. CD45) can have slightly different fluorescence spillover therefore, it may also be necessary to use clone-specific compensation controls when using these reagents.