The 53-5.8 monoclonal antibody specifically binds to the β chain of the CD8 differentiation antigen (Ly-3.2 or Lyt-3.2) of most mouse strains, having weak reactivity with Ly-3.1 strains (e.g., AKR, C58, MRL, PL). The CD8 α and α' chains (CD8a) form heterodimers with the CD8 β chain (CD8b, Ly-3, or Lyt-3) on the surface of most thymocytes. A subpopulation of mature T lymphocytes (i.e., MHC class I-restricted T cells, including most T suppressor/cytotoxic cells) expresses almost exclusively the CD8 αβ heterodimer (the α' chain is absent). Subsets of γδ TCR-bearing T cells, intestinal intraepithelial lymphocytes, and dendritic cells express CD8a without CD8b. It has been suggested that the expression of the CD8a/CD8b heterodimer is restricted to T lymphocytes which matured in the thymus or in an extrathymic environment that had been influenced by thymus-initiated neuroendocrine signals. CD8 is an antigen coreceptor on the T-cell surface which interacts with MHC class I molecules on antigen-presenting cells. It participates in T-cell activation through its association with the T-cell receptor complex and protein tyrosine kinase lck (p56lck).
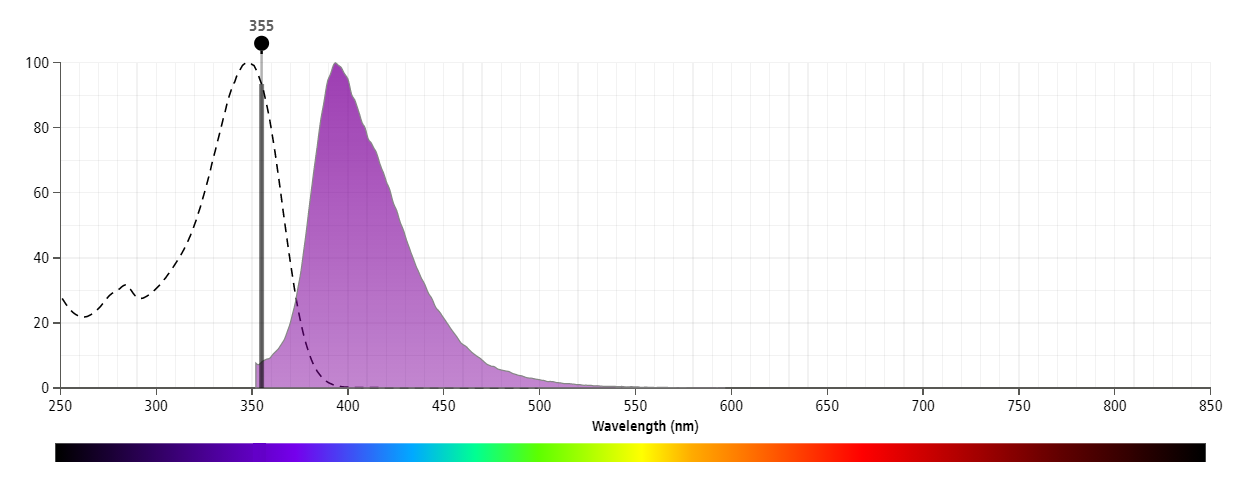
The antibody was conjugated to BD Horizon BUV395 which is part of the BD Horizon Brilliant™ Ultraviolet family of dyes. With an Ex Max near 348 nm and an Em Max near 395 nm, BD Horizon BUV395 can be excited by the ultraviolet laser (355 nm) laser and detected with a 379/28 filter. This dye has been exclusively developed by BD Biosciences as an optimal dye for use on instruments equipped with the ultraviolet laser and has virtually no spillover into any other detector.