
Preparation And Storage
Product Notices
- Since applications vary, each investigator should titrate the reagent to obtain optimal results.
- An isotype control should be used at the same concentration as the antibody of interest.
- Warning: Some APC-Cy7 and PE-Cy7 conjugates show changes in their emission spectrum with prolonged exposure to formaldehyde. If you are unable to analyze fixed samples within four hours, we recommend that you use BD™ Stabilizing Fixative (Cat. No. 338036).
- Please observe the following precautions: Absorption of visible light can significantly alter the energy transfer occurring in any tandem fluorochrome conjugate; therefore, we recommend that special precautions be taken (such as wrapping vials, tubes, or racks in aluminum foil) to prevent exposure of conjugated reagents, including cells stained with those reagents, to room illumination.
- Cy is a trademark of Amersham Biosciences Limited. This conjugated product is sold under license to the following patents: US Patent Nos. 5,486,616; 5,569,587; 5,569,766; 5,627,027.
- This product is subject to proprietary rights of Amersham Biosciences Corp. and Carnegie Mellon University and made and sold under license from Amersham Biosciences Corp. This product is licensed for sale only for research. It is not licensed for any other use. If you require a commercial license to use this product and do not have one return this material, unopened to BD Biosciences, 10975 Torreyana Rd, San Diego, CA 92121 and any money paid for the material will be refunded.
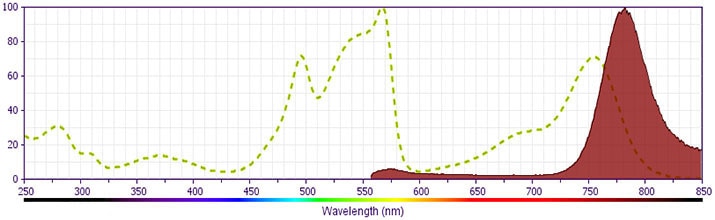
- PE-Cy7 is a tandem fluorochrome composed of R-phycoerythrin (PE), which is excited by 488-nm light and serves as an energy donor, coupled to the cyanine dye Cy7, which acts as an energy acceptor and fluoresces maximally at 780 nm. PE-Cy7 tandem fluorochrome emission is collected in a detector for fluorescence wavelengths of 750 nm and higher. Although every effort is made to minimize the lot-to-lot variation in the efficiency of the fluorochrome energy transfer, differences in the residual emission from PE may be observed. Therefore, we recommend that individual compensation controls be performed for every PE-Cy7 conjugate. PE-Cy7 is optimized for use with a single argon ion laser emitting 488-nm light, and there is no significant overlap between PE-Cy7 and FITC emission spectra. When using dual-laser cytometers, which may directly excite both PE and Cy7, we recommend the use of cross-beam compensation during data acquisition or software compensation during data analysis.
- Caution: Sodium azide yields highly toxic hydrazoic acid under acidic conditions. Dilute azide compounds in running water before discarding to avoid accumulation of potentially explosive deposits in plumbing.
- For fluorochrome spectra and suitable instrument settings, please refer to our Multicolor Flow Cytometry web page at www.bdbiosciences.com/colors.
- Please refer to www.bdbiosciences.com/us/s/resources for technical protocols.
Companion Products

.png?imwidth=320)
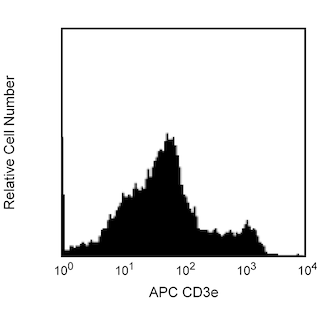
The M1/69 monoclonal antibody specifically binds to CD24 (Heat-Stable Antigen, HSA or HsAg), a variably glycosylated, glycosyl-phosphatidylinositol-anchored membrane protein expressed on erythrocytes, granulocytes, monocytes, lymphocytes, and neurons. Hematopoietic stem cells of the embryonic yolk sac and fetal liver express CD24. Levels of expression of CD24 vary during differentiation of the T and B cell lineages. In the bone marrow, hematopoietic progenitors acquire CD24 expression upon commitment to the B-lymphocyte lineage. Immature B cells in the bone marrow express low CD24 levels whereas peripheral B lymphocytes express intermediate to high levels of CD24. The level of CD24 expression has been reported to rise upon activation of splenic B cells with LPS, but not with CD154 (CD40 Ligand). The majority of thymocytes express high levels of CD24, while most mature thymic and peripheral T lymphocytes do not express CD24. In contrast, TCR-bearing thymocytes which emigrate to the spleen are CD24+. Dendritic cells of the thymus, spleen, liver, and epidermal Langerhans cells have also been reported to express CD24. CD24 is not expressed by NK cells, as determined by staining with J11d mAb (Cat. No. 553146). CD24 is involved in the costimulation of CD4+ T cells by B cells, it is a "co-inducer" of in vitro thymocyte maturation, and it is a ligand of CD62P (P-selectin). While the monoclonal antibodies 30-F1, M1/69, and J11d all react with CD24, they show subtle differences in the level of staining of different lymphocyte populations. When possible, investigators should continue to use the same monoclonal anti-CD24 antibody as used in previous studies.
Development References (23)
-
Aigner S, Ruppert M, Hubbe M, et al. Heat stable antigen (mouse CD24) supports myeloid cell binding to endothelial and platelet P-selectin. Int Immunol. 1995; 7(10):1557-1565. (Biology). View Reference
-
Allman DM, Ferguson SE, Cancro MP. Peripheral B cell maturation. I. Immature peripheral B cells in adults are heat-stable antigenhi and exhibit unique signaling characteristics. J Immunol. 1992; 149(8):2533-2540. (Biology). View Reference
-
Allman DM, Ferguson SE, Lentz VM, Cancro MP. Peripheral B cell maturation. II. Heat-stable antigen(hi) splenic B cells are an immature developmental intermediate in the production of long-lived marrow-derived B cells. J Immunol. 1993; 151(9):4431-4444. (Biology). View Reference
-
Alterman LA, Crispe IN, Kinnon C. Characterization of the murine heat-stable antigen: an hematolymphoid differentiation antigen defined by the J11d, M1/69 and B2A2 antibodies. Eur J Immunol. 1990; 20(7):1597-1602. (Biology). View Reference
-
Ardavin C, Wu L, Ferrero I, Shortman K. Mouse thymic dendritic cell subpopulations. Immunol Lett. 1993; 38(1):19-25. (Biology). View Reference
-
Auerbach R, Huang H, Lu L. Hematopoietic stem cells in the mouse embryonic yolk sac. Stem Cells. 1996; 14(3):269-280. (Biology). View Reference
-
Bruce J, Symington FW, McKearn TJ, Sprent J. A monoclonal antibody discriminating between subsets of T and B cells. J Immunol. 1981; 127(6):2496-2501. (Biology). View Reference
-
Calaora V, Chazal G, Nielsen PJ, Rougon G, Moreau H. mCD24 expression in the developing mouse brain and in zones of secondary neurogenesis in the adult. Neuroscience. 1996; 73(2):581-594. (Biology). View Reference
-
Cibotti R, Punt JA, Dash KS, Sharrow SO, Singer A. Surface molecules that drive T cell development in vitro in the absence of thymic epithelium and in the absence of lineage-specific signals. Immunity. 1997; 6(3):245-255. (Biology). View Reference
-
Crispe IN, Bevan MJ. Expression and functional significance of the J11d marker on mouse thymocytes. J Immunol. 1987 April; 138(7):2013-2018. (Biology). View Reference
-
Crowley M, Inaba K, Witmer-Pack M, Steinman RM. The cell surface of mouse dendritic cells: FACS analyses of dendritic cells from different tissues including thymus. Cell Immunol. 1989; 118(1):108-125. (Biology). View Reference
-
Hardy RR, Carmack CE, Shinton SA, Kemp JD, Hayakawa K. Resolution and characterization of pro-B and pre-pro-B cell stages in normal mouse bone marrow. J Exp Med. 1991; 173(5):1213-1225. (Biology). View Reference
-
Hunte BE, Capone M, Zlotnik A, Rennick D, Moore TA. Acquisition of CD24 expression by Lin-CD43+B220(low)ckit(hi) cells coincides with commitment to the B cell lineage. Eur J Immunol. 1998; 28(11):3850-3856. (Biology). View Reference
-
Kelly KA, Pearse M, Lefrancois L, Scollay R. Emigration of selected subsets of gamma delta + T cells from the adult murine thymus. Int Immunol. 1993; 5(4):331-335. (Biology). View Reference
-
Kennedy MK, Mohler KM, Shanebeck KD, et al. Induction of B cell costimulatory function by recombinant murine CD40 ligand. Eur J Immunol. 1994; 24(1):116-123. (Biology). View Reference
-
Ledbetter JA, Herzenberg LA. Xenogeneic monoclonal antibodies to mouse lymphoid differentiation antigens. Immunol Rev. 1979; 47:63-90. (Biology). View Reference
-
Li YS, Hayakawa K, Hardy RR. The regulated expression of B lineage associated genes during B cell differentiation in bone marrow and fetal liver. J Exp Med. 1993; 178(3):951-960. (Biology). View Reference
-
Springer T, Galfre G, Secher DS, Milstein C. Monoclonal xenogeneic antibodies to murine cell surface antigens: identification of novel leukocyte differentiation antigens. Eur J Immunol. 1978; 8(8):539-551. (Immunogen). View Reference
-
Veillette A, Zuniga-Pflucker JC, Bolen JB, Kruisbeek AM. Engagement of CD4 and CD8 expressed on immature thymocytes induces activation of intracellular tyrosine phosphorylation pathways. J Exp Med. 1989; 170(5):1671-1680. (Biology). View Reference
-
Vremec D, Zorbas M, Scollay R, et al. The surface phenotype of dendritic cells purified from mouse thymus and spleen: investigation of the CD8 expression by a subpopulation of dendritic cells. J Exp Med. 1992; 176(1):47-58. (Biology). View Reference
-
Wenger RH, Rochelle JM, Seldin MF, Kohler G, Nielsen PJ. The heat stable antigen (mouse CD24) gene is differentially regulated but has a housekeeping promoter. J Biol Chem. 1993; 268(31):23345-23352. (Biology). View Reference
-
Wilson A, Day LM, Scollay R, Shortman K. Subpopulations of mature murine thymocytes: properties of CD4-CD8+ and CD4+CD8- thymocytes lacking the heat-stable antigen. Cell Immunol. 1988; 117(2):312-326. (Biology). View Reference
-
Woo J, Lu L, Rao AS, et al. Isolation, phenotype, and allostimulatory activity of mouse liver dendritic cells. Transplantation. 1994; 58(4):484-491. (Biology). View Reference
Please refer to Support Documents for Quality Certificates
Global - Refer to manufacturer's instructions for use and related User Manuals and Technical data sheets before using this products as described
Comparisons, where applicable, are made against older BD Technology, manual methods or are general performance claims. Comparisons are not made against non-BD technologies, unless otherwise noted.
For Research Use Only. Not for use in diagnostic or therapeutic procedures.