






Preparation And Storage
Recommended Assay Procedures
Mouse BD Fc Block™ purified anti-CD16/CD32 mAb 2.4G2 (Cat. No. 553141/553142) may help to reduce non-specific binding of GL1 antibody to cells bearing Fcγ-receptors. When performing immunofluorescent staining in the presence of Mouse BD Fc Block™, a second step antibody which does not cross-react with the 2.4G2 antibody (Rat IgG2b, κ) must be used. We have found that FITC-conjugated mouse anti-rat IgG2a (clone RG7/1.30, Cat. No. 553896) works well for this purpose.
Immunohistochemistry: For IHC, we recommend the use of purified GL1 mAb in our special formulation for immunohistochemistry, Cat. No. 550542).
Product Notices
- Since applications vary, each investigator should titrate the reagent to obtain optimal results.
- Please refer to www.bdbiosciences.com/us/s/resources for technical protocols.
- Caution: Sodium azide yields highly toxic hydrazoic acid under acidic conditions. Dilute azide compounds in running water before discarding to avoid accumulation of potentially explosive deposits in plumbing.
- Sodium azide is a reversible inhibitor of oxidative metabolism; therefore, antibody preparations containing this preservative agent must not be used in cell cultures nor injected into animals. Sodium azide may be removed by washing stained cells or plate-bound antibody or dialyzing soluble antibody in sodium azide-free buffer. Since endotoxin may also affect the results of functional studies, we recommend the NA/LE (No Azide/Low Endotoxin) antibody format, if available, for in vitro and in vivo use.
Data Sheets
Companion Products

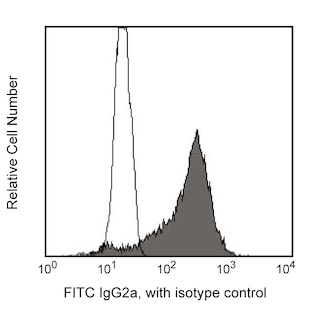
Please refer to Support Documents for Quality Certificates
Global - Refer to manufacturer's instructions for use and related User Manuals and Technical data sheets before using this products as described
Comparisons, where applicable, are made against older BD Technology, manual methods or are general performance claims. Comparisons are not made against non-BD technologies, unless otherwise noted.
Please refer to Support Documents for Quality Certificates
Global - Refer to manufacturer's instructions for use and related User Manuals and Technical data sheets before using this products as described
Comparisons, where applicable, are made against older BD Technology, manual methods or are general performance claims. Comparisons are not made against non-BD technologies, unless otherwise noted.
For Research Use Only. Not for use in diagnostic or therapeutic procedures.