Preparation And Storage
Recommended Assay Procedures
BD® CompBeads can be used as surrogates to assess fluorescence spillover (compensation). When fluorochrome conjugated antibodies are bound to BD® CompBeads, they have spectral properties very similar to cells. However, for some fluorochromes there can be small differences in spectral emissions compared to cells, resulting in spillover values that differ when compared to biological controls. It is strongly recommended that when using a reagent for the first time, users compare the spillover on cells and BD® CompBeads to ensure that BD® CompBeads are appropriate for your specific cellular application.
Product Notices
- When using high concentrations of antibody, background binding of this dye to erythroid fragments produced by ammonium chloride-based lysis, such as with BD Pharm Lyse™ Lysing Buffer (Cat. No. 555899), has been observed when the antibody conjugate was present during the lysis procedure. This may cause nonspecific staining of target cells, such as leukocytes, which have bound the resulting erythroid fragments. This background can be mitigated by any of the following: titrating the antibody conjugate to a lower concentration, fixing samples with formaldehyde, or removing erythrocytes before staining (eg, gradient centrifugation or pre-lysis with wash). This background has not been observed when cells were lysed with BD FACS™ Lysing Solution (Cat. No. 349202) after staining.
- Researchers should determine the optimal concentration of this reagent for their individual applications.
- The production process underwent stringent testing and validation to assure that it generates a high-quality conjugate with consistent performance and specific binding activity. However, verification testing has not been performed on all conjugate lots.
- Please refer to www.bdbiosciences.com/us/s/resources for technical protocols.
- Please observe the following precautions: Absorption of visible light can significantly alter the energy transfer occurring in any tandem fluorochrome conjugate; therefore, we recommend that special precautions be taken (such as wrapping vials, tubes, or racks in aluminum foil) to prevent exposure of conjugated reagents, including cells stained with those reagents, to room illumination.
- An isotype control should be used at the same concentration as the antibody of interest.
- Caution: Sodium azide yields highly toxic hydrazoic acid under acidic conditions. Dilute azide compounds in running water before discarding to avoid accumulation of potentially explosive deposits in plumbing.
- Please refer to http://regdocs.bd.com to access safety data sheets (SDS).
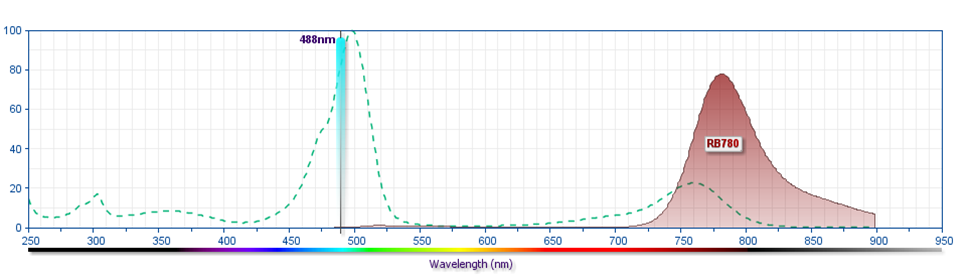
- For fluorochrome spectra and suitable instrument settings, please refer to our Multicolor Flow Cytometry web page at www.bdbiosciences.com/colors.
- Since applications vary, each investigator should titrate the reagent to obtain optimal results.
- Cy is a trademark of Global Life Sciences Solutions Germany GmbH or an affiliate doing business as Cytiva.
Companion Products



The A1 monoclonal antibody specifically binds to the Ly-49A[B6] alloantigen, an inhibitory receptor that is expressed on subsets of natural killer (NK) cells and NK-1.1-positive T lymphocytes (NKT cells) in C57BL/6, C57BL/10, and B10 congenic mice, on a population of memory CD8+ T lymphocytes and NK1.1+ γδ T cells in C57BL/6 mice, and on a distinct subset of B-1 cells (CD5+B220[lo]) of C57BL/6 mice. The A1 antibody has also been reported to crossreact with Ly-49ANOD, Ly-49PNOD, Ly-49P129/J, and Ly-49V129/J alloantigens. The proportion of NKT cells expressing Ly-49A is higher (2-5 fold) in thymus than in liver (immature and mature NKT cells, respectively), and there is evidence that the down regulation of Ly-49 receptor expression is necessary for normal NKT cell development to occur. Most NK cells express a single allele of Ly-49A, although occasionally they may express more than one allele. The Ly-49 family of NK-cell receptors, members of the C-type lectin superfamily, are disulfide-linked type-II transmembrane protein homodimers with extracellular carbohydrate-recognition domains (CRD) that bind to MHC class I alloantigens. The A1 antibody is specific for the Ly-49A[B6] CRD. The Ly-49 family members are expressed independently, such that an individual NK or T cell may display more than one class of Ly-49 receptor homodimers. The Ly-49A[B6] allonantigen binds to H-2D[d], H-2D[k], and H-2D[p], and the A1 antibody blocks this binding. Binding of Ly-49A[B6] to lyphoblasts expressing MHC class I antigens of the f, q, r, s, and v haplotypes has also been demonstrated. The levels of the Ly-49 inhibitory receptors are down-regulated by their ligands in vivo, and various levels of expression of a Ly-49 inhibitory receptor may affect the specificity of NK cells. In vitro studies suggest that the Ly-49A receptor mediates negative regulation of NK-cell cytolytic activity via tyrosine phosphorylation of its ITIM (Immunoreceptor Tyrosine-based Inhibitory Motif).
Development References (31)
-
Bendelac A. Mouse NK1+ T cells. Curr Opin Immunol. 1995; 7(3):367-374. (Clone-specific). View Reference
-
Brennan J, Mahon G, Mager DL, Jefferies WA, Takei F. Recognition of class I major histocompatibility complex molecules by Ly-49: specificities and domain interactions. J Exp Med. 1996; 183(4):1553-1559. (Biology). View Reference
-
Chang CS, Kane KP. Evidence for sulfate modification of H-2Dd on N-linked carbohydrate(s): possible involvement in Ly-49A interaction. J Immunol. 1998; 160(9):4367-4374. (Biology). View Reference
-
Coles MC, McMahon CW, Takizawa H, Raulet DH. Memory CD8 T lymphocytes express inhibitory MHC-specific Ly49 receptors. Eur J Immunol. 2000; 30(1):236-244. (Biology). View Reference
-
Daniels BF, Karlhofer FM, Seaman WE, Yokoyama WM. A natural killer cell receptor specific for a major histocompatibility complex class I molecule. J Exp Med. 1994; 180(2):687-692. (Biology). View Reference
-
Guy-Grand D, Cuenod-Jabri B, Malassis-Seris M, Selz F, Vassalli P. Complexity of the mouse gut T cell immune system: identification of two distinct natural killer T cell intraepithelial lineages. Eur J Immunol. 1996; 26(9):2248-2256. (Clone-specific). View Reference
-
Hanke T, Takizawa H, McMahon CW, et al. Direct assessment of MHC class I binding by seven Ly49 inhibitory NK cell receptors. Immunity. 1999; 11(1):67-77. (Biology). View Reference
-
Hara T, Nishimura H, Hasegawa Y, Yoshikai Y. Thymus-dependent modulation of Ly49 inhibitory receptor expression on NK1.1+gamma/delta T cells. Immunology. 2001; 102(1):24-30. (Biology). View Reference
-
Held W, Kunz B. An allele-specific, stochastic gene expression process controls the expression of multiple Ly49 family genes and generates a diverse, MHC-specific NK cell receptor repertoire. Eur J Immunol. 1998; 28(8):2407-2416. (Biology). View Reference
-
Hoglund P, Sundback J, Olsson-Alheim MY, et al. Host MHC class I gene control of NK-cell specificity in the mouse. Immunol Rev. 1997; 155:11-28. (Biology). View Reference
-
Kane KP. Ly-49 mediates EL4 lymphoma adhesion to isolated class I major histocompatibility complex molecules. J Exp Med. 1994; 179(3):1011-1015. (Biology). View Reference
-
Karlhofer FM, Ribaudo RK, Yokoyama WM. MHC class I alloantigen specificity of Ly-49+ IL-2-activated natural killer cells. Nature. 1992; 358(6381):66-70. (Clone-specific). View Reference
-
Kase A, Johansson MH, Olsson-Alheim MY, Karre K, Hoglund P. External and internal calibration of the MHC class I-specific receptor Ly49A on murine natural killer cells. J Immunol. 1998; 161(11):6133-6138. (Biology). View Reference
-
Kim S, Yokoyama WM. NK cell granule exocytosis and cytokine production inhibited by Ly-49A engagement. Cell Immunol. 1998; 183(2):106-112. (Biology). View Reference
-
Liu J, Yu YY, Lindahl KF, Kumar V, Bennett M. Allorecognition by murine natural killer cells: studies with bone marrow transplants and lysis of T lymphoblasts. Chem Immunol. 1996; 64:164-180. (Biology). View Reference
-
Makrigiannis AP, Pau AT, Saleh A, Winkler-Pickett R, Ortaldo JR, Anderson SK. Class I MHC-binding characteristics of the 129/J Ly49 repertoire. J Immunol. 2001; 166(8):5034-5043. (Biology). View Reference
-
Mason LH, Gosselin P, Anderson SK, Fogler WE, Ortaldo JR, McVicar DW. Differential tyrosine phosphorylation of inhibitory versus activating Ly-49 receptor proteins and their recruitment of SHP-1 phosphatase. J Immunol. 1997; 159(9):4187-4196. (Biology). View Reference
-
Mason LH, Willette-Brown J, Anderson SK, et al. Characterization of an associated 16-kDa tyrosine phosphoprotein required for Ly-49D signal transduction. J Immunol. 1998; 160(9):4148-4152. (Biology). View Reference
-
Matsumoto N, Ribaudo RK, Abastado JP, Margulies DH, Yokoyama WM. The lectin-like NK cell receptor Ly-49A recognizes a carbohydrate-independent epitope on its MHC class I ligand. Immunity. 1998; 8(2):245-254. (Biology). View Reference
-
Nagasawa R, Gross J, Kanagawa O, et al. Identification of a novel T cell surface disulfide-bonded dimer distinct from the alpha/beta antigen receptor. J Immunol. 1987; 138(3):815-824. (Immunogen). View Reference
-
Ochi H, Watanabe T. Negative regulation of B cell receptor-mediated signaling in B-1 cells through CD5 and Ly49 co-receptors via Lyn kinase activity. Int Immunol. 2000; 12(10):1417-1423. (Biology). View Reference
-
Olsson-Alheim MY, Salcedo M, Ljunggren HG, Karre K, Sentman CL. NK cell receptor calibration: effects of MHC class I induction on killing by Ly49Ahigh and Ly49Alow NK cells. J Immunol. 1997; 159(7):3189-3194. (Biology). View Reference
-
Olsson-Alheim MY, Sundback J, Karre K, Sentman CL. The MHC class I molecule H-2Dp inhibits murine NK cells via the inhibitory receptor Ly49A. J Immunol. 1999; 162(12):7010-7014. (Biology). View Reference
-
Raulet DH, Held W, Correa I, Dorfman JR, Wu MF, Corral L. Specificity, tolerance and developmental regulation of natural killer cells defined by expression of class I-specific Ly49 receptors. Immunol Rev. 1997; 155:41-52. (Biology). View Reference
-
Robson MacDonald H, Lees RK, Held W. Developmentally regulated extinction of Ly-49 receptor expression permits maturation and selection of NK1.1+ T cells. J Exp Med. 1998; 187(12):2109-2114. (Biology). View Reference
-
Silver ET, Gong DE, Chang CS, Amrani A, Santamaria P, Kane KP. Ly-49P activates NK-mediated lysis by recognizing H-2Dd. J Immunol. 2000; 165(4):1771-1781. (Biology). View Reference
-
Skold M, Cardell S. Differential regulation of Ly49 expression on CD4+ and CD4-CD8- (double negative) NK1.1+ T cells. Eur J Immunol. 2000; 30(9):2488-2496. (Clone-specific). View Reference
-
Sundback J, Nakamura MC, Waldenstrom M, et al. The alpha2 domain of H-2Dd restricts the allelic specificity of the murine NK cell inhibitory receptor Ly-49A. J Immunol. 1998; 160(12):5971-5978. (Biology). View Reference
-
Yokoyama WM, Kehn PJ, Cohen DI, Shevach EM. Chromosomal location of the Ly-49 (A1, YE1/48) multigene family. Genetic association with the NK 1.1 antigen. J Immunol. 1990; 145(7):2353-2358. (Clone-specific). View Reference
-
Yokoyama WM, Seaman WE. The Ly-49 and NKR-P1 gene families encoding lectin-like receptors on natural killer cells: the NK gene complex. Annu Rev Immunol. 1993; 11:613-635. (Clone-specific). View Reference
-
Yu YY, George T, Dorfman JR, Roland J, Kumar V, Bennett M. The role of Ly49A and 5E6(Ly49C) molecules in hybrid resistance mediated by murine natural killer cells against normal T cell blasts. Immunity. 1996; 4(1):67-76. (Biology). View Reference
Please refer to Support Documents for Quality Certificates
Global - Refer to manufacturer's instructions for use and related User Manuals and Technical data sheets before using this products as described
Comparisons, where applicable, are made against older BD Technology, manual methods or are general performance claims. Comparisons are not made against non-BD technologies, unless otherwise noted.
For Research Use Only. Not for use in diagnostic or therapeutic procedures.