

Preparation And Storage
Recommended Assay Procedures
Recommended Assay Procedure:
Precautions for flow cytometry: For flow cytometry of cell suspensions from peripheral lymphoid tissues, it is recommended that multicolor staining be performed to identify T lymphocytes and/or NK cells and that purified anti-mouse CD16/CD32 mAb 2.4G2 [Mouse BD Fc Block™] (Cat. No. 553141) be used. Since this antigen is expressed at low density on resting peripheral T lymphocytes, it may be desirable to amplify staining by using a biotinylated second-step antibody followed by a "bright" third-step reagent, such as streptavidin-PE (Cat. No. 554061). If Mouse BD Fc Block™ is used, it is important that the second-step anti-hamster IgG antibody does not cross-react with the Mouse BD Fc Block™. Biotinylated anti-Syrian hamster IgG2 mAb G192-3 (Cat. No. 554029) would be a suggested second-step antibody to consider.
Caution: Sodium azide is a reversible inhibitor of oxidative metabolism; therefore, antibody preparations containing this preservative agent must not be used in cell cultures nor injected into animals. Sodium azide may be removed by washing stained cells or plate-bound antibody or dialyzing soluble antibody in sodium azide-free buffer. Since endotoxin may also effect the results of functional studies, we recommend the NA/LE™ (No Azide/Low Endotoxin) antibody format for in vitro and in vivo use.
Product Notices
- Since applications vary, each investigator should titrate the reagent to obtain optimal results.
- Please refer to www.bdbiosciences.com/us/s/resources for technical protocols.
- Although hamster immunoglobulin isotypes have not been well defined, BD Biosciences Pharmingen has grouped Armenian and Syrian hamster IgG monoclonal antibodies according to their reactivity with a panel of mouse anti-hamster IgG mAbs. A table of the hamster IgG groups, Reactivity of Mouse Anti-Hamster Ig mAbs, may be viewed at http://www.bdbiosciences.com/documents/hamster_chart_11x17.pdf.
- Caution: Sodium azide yields highly toxic hydrazoic acid under acidic conditions. Dilute azide compounds in running water before discarding to avoid accumulation of potentially explosive deposits in plumbing.
Companion Products


.png?imwidth=320)
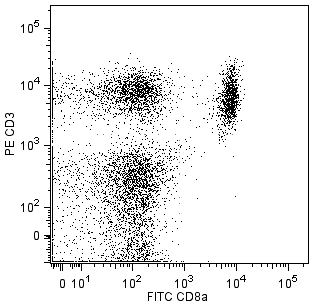
.png?imwidth=320)

The 37.51 antibody reacts with CD28, which is expressed on most thymocytes, at low density on nearly all CD4+ and CD8+ peripheral T cells, and at even lower density on NK cells. The expression of CD28, in splenocytes and thymocytes, has been reported to increase after activation. CD28 transcripts are found in mast cells, and cell-surface expression of CD28 is induced upon maturation or activation of mast cells. It has been reported that CD28 is not expressed on some populations of intraepithelial T lymphocytes. CD28 is a costimulatory receptor; its ligands include CD80 (B7-1) and CD86 (B7-2). The 37.51 mAb augments proliferation and cytokine production by activated T and NK cells and can provide a costimulatory signal for CTL induction. There is considerable evidence that CD28 is a costimulatory receptor involved in many, but not all, T cell-dependent immune responses.
This antibody is routinely tested by flow cytometric analysis. Other applications were tested at BD Biosciences Pharmingen during antibody development only or reported in the literature.
Development References (14)
-
Bluestone JA. New perspectives of CD28-B7-mediated T cell costimulation. Immunity. 1995; 2(6):555-559. (Biology). View Reference
-
Cibotti R, Punt JA, Dash KS, Sharrow SO, Singer A. Surface molecules that drive T cell development in vitro in the absence of thymic epithelium and in the absence of lineage-specific signals. Immunity. 1997; 6(3):245-255. (Biology). View Reference
-
Gelfanov V, Lai YG, Gelfanova V, Dong JY, Su JP, Liao NS. Differential requirement of CD28 costimulation for activation of murine CD8+ intestinal intraepithelial lymphocyte subsets and lymph node cells. J Immunol. 1995; 155(1):76-82. (Biology). View Reference
-
Gross JA, Callas E, Allison JP. Identification and distribution of the costimulatory receptor CD28 in the mouse. J Immunol. 1992; 149(2):380-388. (Immunogen: (Co)-stimulation, Immunoprecipitation). View Reference
-
Harding FA, Allison JP. CD28-B7 interactions allow the induction of CD8+ cytotoxic T lymphocytes in the absence of exogenous help. J Exp Med. 1993; 177(6):1791-1796. (Biology: (Co)-stimulation). View Reference
-
Harding FA, McArthur JG, Gross JA, Raulet DH, Allison JP. CD28-mediated signalling co-stimulates murine T cells and prevents induction of anergy in T-cell clones. Nature. 1992; 356(6370):607-609. (Biology: (Co)-stimulation). View Reference
-
June CH, Bluestone JA, Nadler LM, Thompson CB. The B7 and CD28 receptor families. Immunol Today. 1994; 15(7):321-331. (Biology: (Co)-stimulation). View Reference
-
Krummel MF, Allison JP. CD28 and CTLA-4 have opposing effects on the response of T cells to stimulation. J Exp Med. 1995; 182(2):459-465. (Biology: (Co)-stimulation). View Reference
-
Marietta EV, Weis JJ, Weis JH. CD28 expression by mouse mast cells is modulated by lipopolysaccharide and outer surface protein A lipoprotein from Borrelia burgdorferi. J Immunol. 1997; 159(6):2840-2848. (Biology). View Reference
-
Masten BJ, Yates JL, Pollard Koga AM, Lipscomb MF. Characterization of accessory molecules in murine lung dendritic cell function: roles for CD80, CD86, CD54, and CD40L. Am J Respir Cell Mol Biol. 1997; 16(3):335-342. (Biology: Blocking). View Reference
-
Nandi D, Gross JA, Allison JP. CD28-mediated costimulation is necessary for optimal proliferation of murine NK cells. J Immunol. 1994; 152(7):3361-3369. (Biology: (Co)-stimulation). View Reference
-
Nishio M, Spielman J, Lee RK, Nelson DL, Podack ER. CD80 (B7.1) and CD54 (intracellular adhesion molecule-1) induce target cell susceptibility to promiscuous cytotoxic T cell lysis. J Immunol. 1996; 157(10):4347-4353. (Biology: (Co)-stimulation, Cytotoxicity). View Reference
-
Rakasz E, Hagen M, Sandor M, Lynch RG. Gamma delta T cells of the murine vagina: T cell response in vivo in the absence of the expression of CD2 and CD28 molecules. Int Immunol. 1997; 9(1):161-167. (Biology). View Reference
-
Shahinian A, Pfeffer K, Lee KP, et al. Differential T cell costimulatory requirements in CD28-deficient mice. Science. 1993; 261(5121):609-612. (Biology). View Reference
Please refer to Support Documents for Quality Certificates
Global - Refer to manufacturer's instructions for use and related User Manuals and Technical data sheets before using this products as described
Comparisons, where applicable, are made against older BD Technology, manual methods or are general performance claims. Comparisons are not made against non-BD technologies, unless otherwise noted.
For Research Use Only. Not for use in diagnostic or therapeutic procedures.