

Preparation And Storage
Product Notices
- An isotype control should be used at the same concentration as the antibody of interest.
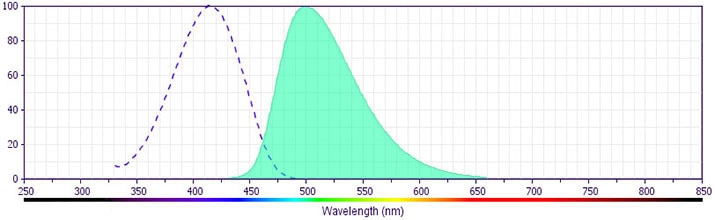
- BD Horizon V500 has a maximum absorption of 415 nm and maximum emission of 500 nm. Before staining with this reagent, please confirm that your flow cytometer is capable of exciting the fluorochrome and discriminating the resulting fluorescence.
- BD Horizon V500 is covered by one or more of the following US patents: 8,431,416.
- Caution: Sodium azide yields highly toxic hydrazoic acid under acidic conditions. Dilute azide compounds in running water before discarding to avoid accumulation of potentially explosive deposits in plumbing.
- For fluorochrome spectra and suitable instrument settings, please refer to our Multicolor Flow Cytometry web page at www.bdbiosciences.com/colors.
- Please refer to www.bdbiosciences.com/us/s/resources for technical protocols.
- Since applications vary, each investigator should titrate the reagent to obtain optimal results.
Companion Products


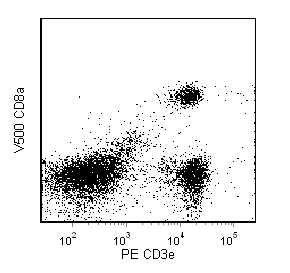
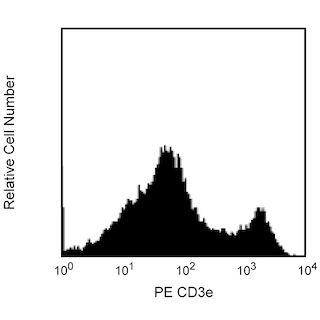
The 53-6.7 monoclonal antibody specifically binds to the 38 kDa α and 34 kDa α' chains of the CD8 differentiation antigen (Ly-2 or Lyt-2) of all mouse strains tested. The CD8 α and α' chains (CD8a) form heterodimers with the CD8 β chain (CD8b, Ly-3, or Lyt-3) on the surface of most thymocytes. A subpopulation of mature T lymphocytes (i.e., MHC class I-restricted T cells, including most T suppressor/cytotoxic cells) expresses almost exclusively the CD8 αβ heterodimer. Subsets of γδ TCR-bearing T cells, intestinal intrapithelial lymphocytes, and dendritic cells express CD8a without CD8b. It has been suggested that the expression of the CD8a/CD8b heterodimer is restricted to T lymphocytes which matured in the thymus or in an extrathymic environment that had been influenced by thymus-initiated neuroendocrine signals. CD8 is an antigen coreceptor on the T-cell surface which interacts with MHC class I molecules on antigen-presenting cells or epithelial cells. It participates in T-cell activation through its association with the T-cell receptor complex and protein tyrosine kinase lck (p56 [lck]). The CD8 α and α' chains arise from alternatively spliced messengers of a single CD8a gene. The longer α form associates with p56 [lck] via a CXCP motif in its cytoplasmic domain, which it shares with CD4, but not with CD8b. The truncated α' chain is unable to associate with p56 [lck], and it may function to attenuate the CD8-mediated costimulatory signal during intrathymic T-cell maturation. In vivo and in vitro treatment with 53-6.7 mAb has reportedly been effective at depleting CD8+ peripheral T lymphocytes. The 53-6.7 antibody has also been reported to cross-react with CD8 α- and α'-like polypeptides on subsets of thymic and peripheral lymphocytes in the Egyptian toad, Bufo regularis.
The antibody is conjugated to BD Horizon™ V500, which has been developed for use in multicolor flow cytometry experiments and is
available exclusively from BD Biosciences. It is excited by the Violet laser with an Ex max of 415 nm and Em Max at 500 nm. BD Horizon V500 conjugates emit at a similar wavelength to Amcyan yet exhibit reduced spillover into the FITC channel. For more information on BD Horizon V500, visit bdbiosciences.com/colors.
When compensating dyes in this spectral range (such as Horizon™ V500 and AmCyan), the most accurate compensation can be obtained using single stained cellular controls. Due to spectral differences between cells and beads in this channel, using BD CompBeads can result in spillover errors for V500 and AmCyan reagents. Therefore, the use of BD CompBeads or BD CompBeads Plus to determine spillover values for these reagents is not recommended. Different V500 reagents (e.g. CD4 vs. CD45) can have slightly different fluorescence spillover therefore, it may also be necessary to use clone specific compensation controls when using these reagents.
Development References (25)
-
Bierer BE, Sleckman BP, Ratnofsky SE, Burakoff SJ. The biologic roles of CD2, CD4, and CD8 in T-cell activation. Annu Rev Immunol. 1989; 7:579-599. (Biology). View Reference
-
Fujiura Y, Kawaguchi M, Kondo Y, et al. Development of CD8 alpha alpha+ intestinal intraepithelial T cells in beta 2-microglobulin- and/or TAP1-deficient mice. J Immunol. 1996; 156(8):2710-2715. (Biology). View Reference
-
Hathcock KS. T cell depletion by cytotoxic elimination. Curr Protoc Immunol. 1991; 1:3.4.1-3.4.3. (Biology). View Reference
-
Janeway CA Jr. The T cell receptor as a multicomponent signalling machine: CD4/CD8 coreceptors and CD45 in T cell activation. Annu Rev Immunol. 1992; 10:645-674. (Biology). View Reference
-
Kruisbeek AM, Shevach EM. Proliferative assays for T cell function. Curr Protoc Immunol. 2004; 3:3.12.1-3.12.14. (Biology). View Reference
-
LeFrancois L. Extrathymic differentiation of intraepithelial lymphocytes: generation of a separate and unequal T-cell repertoire. Immunol Today. 1991; 12(12):436-438. (Biology). View Reference
-
Ledbetter JA, Herzenberg LA. Xenogeneic monoclonal antibodies to mouse lymphoid differentiation antigens. Immunol Rev. 1979; 47:63-90. (Immunogen). View Reference
-
Ledbetter JA, Rouse RV, Micklem HS, Herzenberg LA. T cell subsets defined by expression of Lyt-1,2,3 and Thy-1 antigens. Two-parameter immunofluorescence and cytotoxicity analysis with monoclonal antibodies modifies current views. J Exp Med. 1980; 152(2):280-295. (Biology). View Reference
-
Ledbetter JA, Seaman WE, Tsu TT, Herzenberg LA. Lyt-2 and lyt-3 antigens are on two different polypeptide subunits linked by disulfide bonds. Relationship of subunits to T cell cytolytic activity. J Exp Med. 1981; 153(6):1503-1516. (Biology). View Reference
-
Leishman AJ, Naidenko OV, Attinger A, et al. T cell responses modulated through interaction between CD8alphaalpha and the nonclassical MHC class I molecule, TL. Science. 2001; 294(5548):1848-1849. (Biology). View Reference
-
MacDonald HR, Schreyer M, Howe RC, Bron C. Selective expression of CD8 alpha (Ly-2) subunit on activated thymic gamma/delta cells. Eur J Immunol. 1990; 20(4):927-930. (Biology). View Reference
-
Mitnacht R, Bischof A, Torres-Nagel N, Hunig T. Opposite CD4/CD8 lineage decisions of CD4+8+ mouse and rat thymocytes to equivalent triggering signals: correlation with thymic expression of a truncated CD8 alpha chain in mice but not rats. J Immunol. 1998; 160(2):700-707. (Biology). View Reference
-
Murosaki S, Yoshikai Y, Ishida A, et al. Failure of T cell receptor V beta negative selection in murine intestinal intra-epithelial lymphocytes. Int Immunol. 1991; 3(10):1005-1013. (Biology). View Reference
-
Nakayama K, Nakayama K, Negishi I, et al. Requirement for CD8 beta chain in positive selection of CD8-lineage T cells. Science. 1994; 263(5150):1131-1133. (Biology). View Reference
-
Negm HI, Mansour MH, Saad AH, Abdel Halim RS. Structural characterization of an Lyt-2/3 homolog expressed on Bufo regularis lymphocytes. Comp Biochem Physiol B Biochem Mol Biol. 1996; 113(1):79-87. (Biology). View Reference
-
O'Rourke AM, Mescher MF. The roles of CD8 in cytotoxic T lymphocyte function. Immunol Today. 1993; 14(4):183-188. (Biology). View Reference
-
Rocha B, Vassalli P, Guy-Grand D. The extrathymic T-cell development pathway. Immunol Today. 1992; 14(3):140-141. (Biology). View Reference
-
Sydora BC, Brossay L, Hagenbaugh A, Kronenberg M, Cheroutre H. TAP-independent selection of CD8+ intestinal intraepithelial lymphocytes. J Immunol. 1996; 156(11):4209-4216. (Biology). View Reference
-
Traver D, Akashi K, Manz M, et al. Development of CD8alpha-positive dendritic cells from a common myeloid progenitor. Science. 2000; 290(5499):2152-2154. (Biology). View Reference
-
Walker ID, Murray BJ, Hogarth PM, Kelso A, McKenzie IF. Comparison of thymic and peripheral T cell Ly-2/3 antigens. Eur J Immunol. 1984; 14(10):906-910. (Biology). View Reference
-
Wang J, Klein JR. Thymus-neuroendocrine interactions in extrathymic T cell development. Science. 1994; 265(5180):1860-1862. (Biology). View Reference
-
Zamoyska R, Derham P, Gorman SD, et al. Inability of CD8 alpha' polypeptides to associate with p56lck correlates with impaired function in vitro and lack of expression in vivo. Nature. 1989; 342(6247):278-281. (Biology). View Reference
-
Zamoyska R, Vollmer AC, Sizer KC, Liaw CW, Parnes JR. Two Lyt-2 polypeptides arise from a single gene by alternative splicing patterns of mRNA. Cell. 1985; 43(1):153-163. (Biology). View Reference
-
Zamoyska R. The CD8 coreceptor revisited: one chain good, two chains better. Immunity. 1994; 1(4):243-246. (Biology). View Reference
-
van Ewijk W, van Soest PL, van den Engh GJ. Fluorescence analysis and anatomic distribution of mouse T lymphocyte subsets defined by monoclonal antibodies to the antigens Thy-1, Lyt-1, Lyt-2, and T-200. J Immunol. 1981; 127(6):2594-2604. (Biology). View Reference
Please refer to Support Documents for Quality Certificates
Global - Refer to manufacturer's instructions for use and related User Manuals and Technical data sheets before using this products as described
Comparisons, where applicable, are made against older BD Technology, manual methods or are general performance claims. Comparisons are not made against non-BD technologies, unless otherwise noted.
For Research Use Only. Not for use in diagnostic or therapeutic procedures.