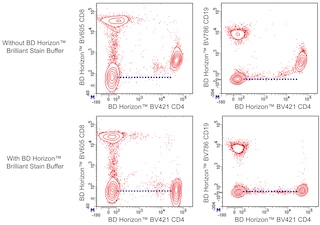
The IM7 antibody specifically recognizes an epitope on both alloantigens and all isoforms of the CD44 glycoprotein (Pgp-1, Ly-24). The standard form of CD44, lacking variable exons and referred to as CD44H or CD44s, is widely expressed on hematopoietic and non-hematopoietic cells. CD44 isoforms encoded by variable exons are expressed on epithelial cells, but only at low levels on most leukocytes. Mice with the Ly-24.1 alloantigen (e.g., BALB/c, CBA/J, DBA/1, DBA/2) have relatively large subsets of CD44H+ T lymphocytes, while Ly-24.2 strains (e.g., A, AKR, CBA/N, C3H/He, C57BL, C57BR, C57L, C58, NZB, SJL, SWR, 129) have fewer CD44H+ T cells. CD44 is a cell adhesion receptor, and its principal ligand, hyaluronate, is a common component of extracellular matrices. Differential glycosylation of CD44 influences its binding to hyaluronate. Additional ligands include the cell surface form of CD74 and the cytokine osteopontin (Eta-1). Bone marrow- and thymus-derived progenitor cells capable of repopulating the thymus express CD44. In the periphery, the level of CD44 expression increases upon activation of B lymphocytes, CD4+ T cells, and CD8+ T cells; memory cells can be recognized by their CD44[hi] phenotype. The IM7 mAb inhibits established collagen-induced arthritis in DBA/1 mice. Moreover, it prevents CNS inflammation and clinical symptoms of experimental autoimmune encephalomyelitis. In contrast, the same antibody exacerbates experimental autoimmune thyroiditis in CBA/J mice. The IM7 mAb recognizes a different epitope from that recognized by mAb KM114, and the antibody pair can be used in ELISA to detect soluble CD44. It has been observed that IM7 antibody crossreacts with human, dog, cat, horse, cow, and pig leukocytes. Anti-human CD44, clone G44-26, and IM7 antibody compete for binding to human peripheral blood lymphocytes.
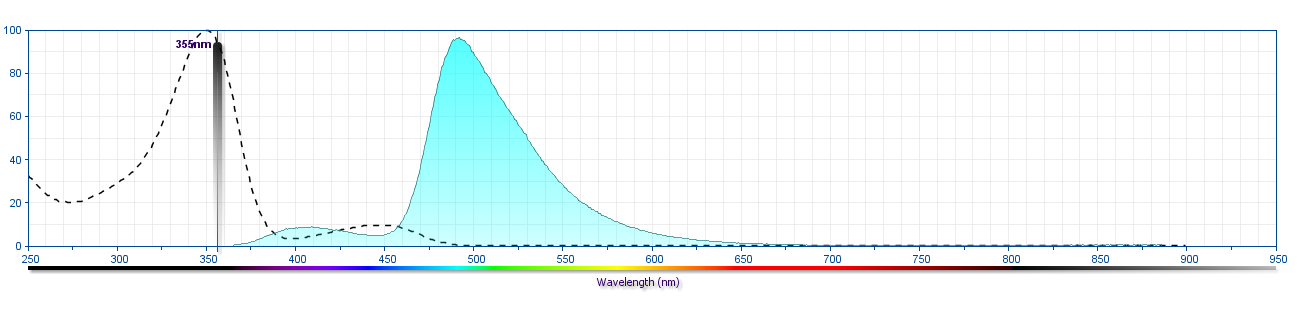
The antibody was conjugated to BD Horizon™ BUV496 which is part of the BD Horizon Brilliant™ Ultraviolet family of dyes. This dye is a tandem fluorochrome of BD Horizon BUV395 with an Ex Max of 348-nm and an acceptor dye with an Em Max at 496-nm. BD Horizon BUV496 can be excited by the ultraviolet laser (355 nm) and detected with a 515/30 nm filter with a 450LP. Due to the excitation of the acceptor dye by other laser lines, there may be significant spillover into the channel detecting BD Horizon V500 or BV510 (eg, 525/40-nm filter). However, the spillover can be corrected through compensation as with any other dye combination.