The A1 monoclonal antibody specifically binds to the Ly-49A[B6] alloantigen, an inhibitory receptor that is expressed on subsets of natural killer (NK) cells and NK-1.1-positive T lymphocytes (NKT cells) in C57BL/6, C57BL/10, and B10 congenic mice, on a population of memory CD8+ T lymphocytes and NK1.1+ γδ T cells in C57BL/6 mice, and on a distinct subset of B-1 cells (CD5+B220[lo]) of C57BL/6 mice. The A1 antibody has also been reported to crossreact with Ly-49ANOD, Ly-49PNOD, Ly-49P129/J, and Ly-49V129/J alloantigens. The proportion of NKT cells expressing Ly-49A is higher (2-5 fold) in thymus than in liver (immature and mature NKT cells, respectively), and there is evidence that the down regulation of Ly-49 receptor expression is necessary for normal NKT cell development to occur. Most NK cells express a single allele of Ly-49A, although occasionally they may express more than one allele. The Ly-49 family of NK-cell receptors, members of the C-type lectin superfamily, are disulfide-linked type-II transmembrane protein homodimers with extracellular carbohydrate-recognition domains (CRD) that bind to MHC class I alloantigens. The A1 antibody is specific for the Ly-49A[B6] CRD. The Ly-49 family members are expressed independently, such that an individual NK or T cell may display more than one class of Ly-49 receptor homodimers. The Ly-49A[B6] allonantigen binds to H-2D[d], H-2D[k], and H-2D[p], and the A1 antibody blocks this binding. Binding of Ly-49A[B6] to lyphoblasts expressing MHC class I antigens of the f, q, r, s, and v haplotypes has also been demonstrated. The levels of the Ly-49 inhibitory receptors are down-regulated by their ligands in vivo, and various levels of expression of a Ly-49 inhibitory receptor may affect the specificity of NK cells. In vitro studies suggest that the Ly-49A receptor mediates negative regulation of NK-cell cytolytic activity via tyrosine phosphorylation of its ITIM (Immunoreceptor Tyrosine-based Inhibitory Motif).
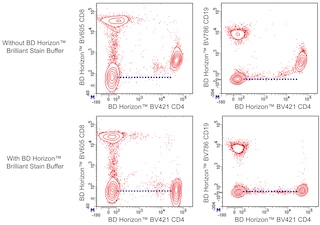
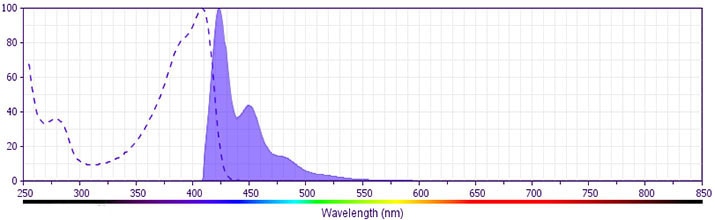
The antibody was conjugated to BD Horizon™ BV421 which is part of the BD Horizon Brilliant™ Violet family of dyes. With an Ex Max of 407-nm and Em Max at 421-nm, BD Horizon BV421 can be excited by the violet laser and detected in the standard Pacific Blue™ filter set (eg, 450/50-nm filter). BD Horizon BV421 conjugates are very bright, often exhibiting a 10 fold improvement in brightness compared to Pacific Blue conjugates.