
Preparation And Storage
Recommended Assay Procedures
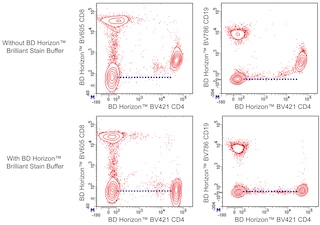
For optimal and reproducible results, BD Horizon Brilliant Stain Buffer should be used anytime two or more BD Horizon Brilliant dyes (including BD OptiBuild Brilliant reagents) are used in the same experiment. Fluorescent dye interactions may cause staining artifacts which may affect data interpretation. The BD Horizon Brilliant Stain Buffer was designed to minimize these interactions. More information can be found in the Technical Data Sheet of the BD Horizon Brilliant Stain Buffer (Cat. No. 563794).
Product Notices
- This antibody was developed for use in flow cytometry.
- The production process underwent stringent testing and validation to assure that it generates a high-quality conjugate with consistent performance and specific binding activity. However, verification testing has not been performed on all conjugate lots.
- Researchers should determine the optimal concentration of this reagent for their individual applications.
- An isotype control should be used at the same concentration as the antibody of interest.
- Caution: Sodium azide yields highly toxic hydrazoic acid under acidic conditions. Dilute azide compounds in running water before discarding to avoid accumulation of potentially explosive deposits in plumbing.
- For fluorochrome spectra and suitable instrument settings, please refer to our Multicolor Flow Cytometry web page at www.bdbiosciences.com/colors.
- Please refer to www.bdbiosciences.com/us/s/resources for technical protocols.
- BD Horizon Brilliant Stain Buffer is covered by one or more of the following US patents: 8,110,673; 8,158,444; 8,575,303; 8,354,239.
Companion Products
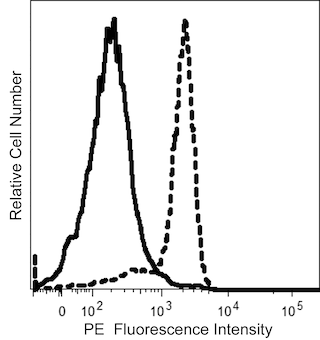



The 11A9 monoclonal antibody specifically binds to CD196, which is also known as CCR6. CCR6 is a seven-transmembrane, G-protein-coupled, glycoprotein receptor that is a member of the beta chemokine receptor family. The human CCR6 gene has been mapped to chromosome 6q27. CCR6 is a receptor for the CC chemokine CCL20/MIP-3alpha/LARC/Exodus and also binds with lower affinity to and mediates responses to beta-defensin2/hBD-2. CCR6 is predominantly expressed by B lymphocytes, certain subsets of effector and memory T cells and by immature dendritic cells but not by monocytes, NK cells, or granulocytes. Skin-homing CLA (Cutaneous Lymphocyte Antigen)-positive memory T cells, Th1 cells, regulatory T cells and IL-17A-producing Th17 cells predominantly express high levels of CCR6. CCR6 mediates the trafficking of T, B, and dendritic cells to epithelial sites near the skin and mucosal surfaces during inflammatory and immunological responses. An N-terminal peptide of human CCR6 was used as an immunogen to generate the 11A9 hybridoma. The 11A9 antibody does not cross-react with human CCR1, CCR2, CCR3, CCR4, CCR5, CCR7, CCR8, CCR9, CXCR1, CXCR2, CXCR3, CXCR4 and CXCR5 receptors. This antibody is NOT a neutralizing antibody.
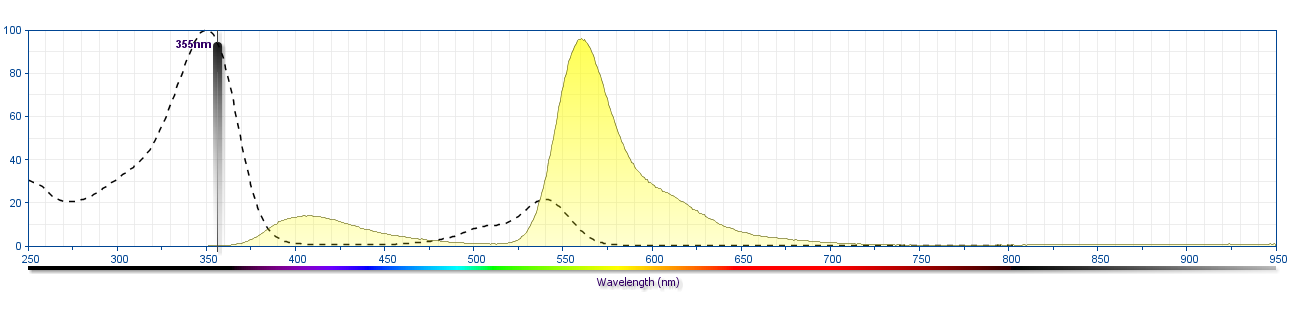
The antibody was conjugated to BD Horizon™ BUV563 which is part of the BD Horizon Brilliant™ Ultraviolet family of dyes. This dye is a tandem fluorochrome of BD Horizon BUV395 which has an Ex Max of 348 nm and an acceptor dye. The tandem has an Em Max at 563 nm. BD Horizon BUV563 can be excited by the 355 nm ultraviolet laser. On instruments with a 561 nm Yellow-Green laser, the recommended bandpass filter is 585/15 nm with a 535 nm long pass to minimize laser light leakage. When BD Horizon BUV563 is used with an instrument that does not have a 561 nm laser, a 560/40 nm filter with a 535 nm long pass may be more optimal. Due to the excitation and emission characteristics of the acceptor dye, there may be spillover into the PE and PE-CF594 detectors. However, the spillover can be corrected through compensation as with any other dye combination.
Development References (15)
-
Baba M, Imai T, Nishimura M, et al. Identification of CCR6, the specific receptor for a novel lymphocyte-directed CC chemokine LARC. J Biol Chem. 1997; 272(23):14893-14898. (Biology). View Reference
-
Brandes M, Willimann K, Lang AB, et al. Flexible migration program regulates gamma delta T-cell involvement in humoral immunity. Blood. 2003; 102(10):3693-3701. (Clone-specific). View Reference
-
Cabezon R, Sintes J, Llinas L, Benitez-Ribas D. Analysis of HLDA9 mAbs on plasmacytoid dendritic cell. Immunol Lett. 2011; 134(2):167-173. (Clone-specific). View Reference
-
Greaves DR, Wang W, Dairaghi DJ, et al. CCR6, a CC chemokine receptor that interacts with macrophage inflammatory protein 3alpha and is highly expressed in human dendritic cells. J Exp Med. 1997; 186(6):837-844. (Biology). View Reference
-
Homey B, Dieu-Nosjean MC, Wiesenborn A, et al. Up-regulation of macrophage inflammatory protein-3 alpha/CCL20 and CC chemokine receptor 6 in psoriasis. J Immunol. 2000; 164(12):6621-6632. (Biology). View Reference
-
Kim CH, Rott L, Kunkel EJ, et al. Rules of chemokine receptor association with T cell polarization in vivo. J Clin Invest. 2001; 108(9):1331-1339. (Biology). View Reference
-
Liao F, Alderson R, Su J, Ullrich SJ, Kreider BL, Farber JM. STRL22 is a receptor for the CC chemokine MIP-3alpha. Biochem Biophys Res Commun. 1997; 236(1):212-217. (Biology). View Reference
-
Liao F, Rabin RL, Smith CS, Sharma G, Nutman TB, Farber JM. CC-chemokine receptor 6 is expressed on diverse memory subsets of T cells and determines responsiveness to macrophage inflammatory protein 3 alpha. J Immunol. 1999; 162(1):186-194. (Biology). View Reference
-
Liao F, Shirakawa AK, Foley JF, Rabin RL, Farber JM. Human B cells become highly responsive to macrophage-inflammatory protein-3 alpha/CC chemokine ligand-20 after cellular activation without changes in CCR6 expression or ligand binding. J Immunol. 2002; 168(10):4871-4880. (Clone-specific). View Reference
-
Lim HW, Lee J, Hillsamer P, Kim CH. Human Th17 cells share major trafficking receptors with both polarized effector T cells and FOXP3+ regulatory T cells. J Immunol. 2008; 180(1):122-129. (Biology). View Reference
-
Llinas L, Lazaro A, de Salort J, Matesanz-Isabel J, Sintes J, Engel P. Expression profiles of novel cell surface molecules on B-cell subsets and plasma cells as analyzed by flow cytometry. Immunol Lett. 2011; 134(2):113-121. (Clone-specific). View Reference
-
Ramos-Medina R, Montes-Moreno S, Maestre L, et al. Immunohistochemical analysis of HLDA9 Workshop antibodies against cell-surface molecules in reactive and neoplastic lymphoid tissues. Immunol Lett. 2011; 134(2):150-156. (Clone-specific). View Reference
-
Sallusto F, Lenig D, Forster R, Lipp M, Lanzavecchia A. Two subsets of memory T lymphocytes with distinct homing potentials and effector functions. Nature. 1999; 401(6754):708-712. (Clone-specific). View Reference
-
Thomas SY, Banerji A, Medoff BD, Lilly CM, Luster AD. Multiple chemokine receptors, including CCR6 and CXCR3, regulate antigen-induced T cell homing to the human asthmatic airway. J Immunol. 2007; 179(3):1901-1912. (Clone-specific). View Reference
-
Yang D, Chertov O, Bykovskaia SN, et al. Beta-defensins: linking innate and adaptive immunity through dendritic and T cell CCR6. Science. 1999; 286(5439):525-528. (Biology). View Reference
Please refer to Support Documents for Quality Certificates
Global - Refer to manufacturer's instructions for use and related User Manuals and Technical data sheets before using this products as described
Comparisons, where applicable, are made against older BD Technology, manual methods or are general performance claims. Comparisons are not made against non-BD technologies, unless otherwise noted.
For Research Use Only. Not for use in diagnostic or therapeutic procedures.