
Preparation And Storage
Product Notices
- Since applications vary, each investigator should titrate the reagent to obtain optimal results.
- This product is subject to proprietary rights of Amersham Biosciences Corp. and Carnegie Mellon University and made and sold under license from Amersham Biosciences Corp. This product is licensed for sale only for research. It is not licensed for any other use. If you require a commercial license to use this product and do not have one return this material, unopened to BD Biosciences, 10975 Torreyana Rd, San Diego, CA 92121 and any money paid for the material will be refunded.
- Cy is a trademark of Amersham Biosciences Limited. This conjugated product is sold under license to the following patents: US Patent Nos. 5,486,616; 5,569,587; 5,569,766; 5,627,027.
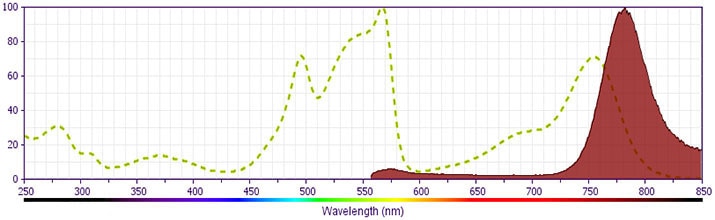
- PE-Cy7 is a tandem fluorochrome composed of R-phycoerythrin (PE), which is excited by 488-nm light and serves as an energy donor, coupled to the cyanine dye Cy7, which acts as an energy acceptor and fluoresces maximally at 780 nm. PE-Cy7 tandem fluorochrome emission is collected in a detector for fluorescence wavelengths of 750 nm and higher. Although every effort is made to minimize the lot-to-lot variation in the efficiency of the fluorochrome energy transfer, differences in the residual emission from PE may be observed. Therefore, we recommend that individual compensation controls be performed for every PE-Cy7 conjugate. PE-Cy7 is optimized for use with a single argon ion laser emitting 488-nm light, and there is no significant overlap between PE-Cy7 and FITC emission spectra. When using dual-laser cytometers, which may directly excite both PE and Cy7, we recommend the use of cross-beam compensation during data acquisition or software compensation during data analysis.
- Caution: Sodium azide yields highly toxic hydrazoic acid under acidic conditions. Dilute azide compounds in running water before discarding to avoid accumulation of potentially explosive deposits in plumbing.
- For fluorochrome spectra and suitable instrument settings, please refer to our Multicolor Flow Cytometry web page at www.bdbiosciences.com/colors.
- Please refer to www.bdbiosciences.com/us/s/resources for technical protocols.
Companion Products

.png?imwidth=320)
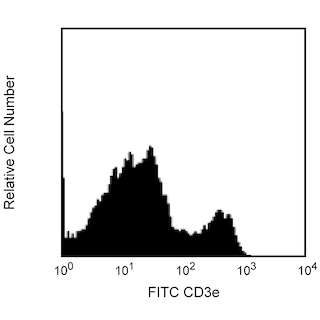
The 1D3 antibody reacts with CD19, a B lymphocyte-lineage differentiation antigen. CD19, a 95-kDa transmembrance glycoprotein, is a member of the immunoglobulin superfamily and is expressed throughout B-lymphocyte development from the pro-B cell through the mature B-cell stages. Terminally differentiated plasma cells do not express CD19. On the surface of mature B cells, the CD19 molecule associates with CD21 (CR-2) and CD81 (TAPA-1), and this multimolecular complex synergizes with surface immunoglobulin to promote cellular activation. Studies with CD19-deficient mice have suggested that the level of CD19 expression affects the generation and maturation of B cells in the bone marrow and periphery. B-1 lineage B cells, also known as CD5+ B cells, are drastically reduced or absent in CD19-deficient mice. Increased levels of CD19 expression correlate with increased frequencies of peritonal and splenic B-1 cells and reduced numbers of conventional B lymphocytes in the periphery. CD19 participates in B-lymphocyte development, B-cell activation, maturation of memory B cells and regulation of tolerance. CD19 has also been detected on peritoneal mast cells, co-localized with CD21/CD35, and it is proposed to play a role in complement-mediated mast-cell activation.
Development References (13)
-
Engel P, Zhou LJ, Ord DC, Sato S, Koller B, Tedder TF. Abnormal B lymphocyte development, activation, and differentiation in mice that lack or overexpress the CD19 signal transduction molecule. Immunity. 1995; 3(1):39-50. (Biology). View Reference
-
Fearon DT. The CD19-CR2-TAPA-1 complex, CD45 and signaling by the antigen receptor of B lymphocytes. Curr Opin Immunol. 1993; 5(3):341-348. (Biology). View Reference
-
Gommerman JL, Oh DY, Zhou X, et al. A role for CD21/CD35 and CD19 in responses to acute septic peritonitis: a potential mechanism for mast cell activation. J Immunol. 2000; 165(12):6915-6921. (Biology). View Reference
-
Inaoki M, Sato S, Weintraub BC, Goodnow CC, Tedder TF. CD19-regulated signaling thresholds control peripheral tolerance and autoantibody production in B lymphocytes. J Exp Med. 1997; 186(11):1923-1931. (Biology). View Reference
-
Krop I, Shaffer AL, Fearon DT, Schlissel MS. The signaling activity of murine CD19 is regulated during cell development. J Immunol. 1996; 157(1):48-56. (Biology: Functional assay). View Reference
-
Krop I, de Fougerolles AR, Hardy RR, Allison M, Schlissel MS, Fearon DT. Self-renewal of B-1 lymphocytes is dependent on CD19. Eur J Immunol. 1996; 26(1):238-242. (Immunogen: Functional assay, Immunoprecipitation). View Reference
-
Rickert RC, Rajewsky K, Roes J. Impairment of T-cell-dependent B-cell responses and B-1 cell development in CD19-deficient mice. Nature. 1995; 376(6538):352-355. (Biology). View Reference
-
Roederer M, Kantor AB, Parks DR, Herzenberg LA. Cy7PE and Cy7APC: bright new probes for immunofluorescence. Cytometry. 1996; 24(3):191-197. (Methodology: Flow cytometry). View Reference
-
Sato S, Jansen PJ, Tedder TF. CD19 and CD22 expression reciprocally regulates tyrosine phosphorylation of Vav protein during B lymphocyte signaling. Proc Natl Acad Sci U S A. 1997; 94(24):13158-13162. (Biology). View Reference
-
Sato S, Miller AS, Howard MC, Tedder TF. Regulation of B lymphocyte development and activation by the CD19/CD21/CD81/Leu 13 complex requires the cytoplasmic domain of CD19. J Immunol. 1997; 159(7):3278-3287. (Biology). View Reference
-
Sato S, Ono N, Steeber DA, Pisetsky DS, Tedder TF. CD19 regulates B lymphocyte signaling thresholds critical for the development of B-1 lineage cells and autoimmunity. J Immunol. 1996; 157(10):4371-4378. (Biology). View Reference
-
Sato S, Steeber DA,Jansen PJ, Tedder TF. CD19 expression levels regulate B lymphocyte development: human CD19 restores normal function in mice lacking endogenous CD19. J Immunol. 1997; 158(10):4662-4669. (Biology). View Reference
-
Tedder TF, Zhou LJ, Engel P. The CD19/CD21 signal transduction complex of B lymphocytes. Immunol Today. 1994; 15(9):437-442. (Biology). View Reference
Please refer to Support Documents for Quality Certificates
Global - Refer to manufacturer's instructions for use and related User Manuals and Technical data sheets before using this products as described
Comparisons, where applicable, are made against older BD Technology, manual methods or are general performance claims. Comparisons are not made against non-BD technologies, unless otherwise noted.
For Research Use Only. Not for use in diagnostic or therapeutic procedures.