The GL1 antibody specifically recognizes the B7-2 (CD86) costimulatory molecule expressed on a broad spectrum of leukocytes, including B lymphocytes, T lymphocytes, thioglycollate-induced peritoneal macrophages, dendritic cells and astrocytes. CD86 is expressed at low levels by freshly explanted peripheral B and T cells, and its expression is substantially increased by a variety of T cell- and B cell-specific stimuli with a peak expression after 18-42 hours of culture. In contrast to most naive CD4+ T cells, memory CD4+ T cells express B7-2, both at the mRNA and protein level. CD86, a ligand for CD28 and CD152 (CTLA-4), is one of the accessory molecules that plays an important role in T cell-B cell costimulatory interactions. It has been shown to be involved in immunoglobulin class-switching and triggering of mouse NK cell-mediated cytotoxicity. CD80 (B7-1) is an alternate ligand for CD28 and CD152 (CTLA-4). GL1 antibody reportedly blocks MLR and stimulation of T cells by natural antigen-presenting cells. In addition, a mixture of anti-B7-1 and anti B7-2 (GL1) mAbs reportedly inhibits the in vitro interaction of CTLA-4 with its ligand and the in vivo priming of cytotoxic T lymphocytes.
The antibody was conjugated to BD Horizon™ BUV661 which is part of the BD Horizon Brilliant™ Ultraviolet family of dyes. This dye is a tandem fluorochrome of BD Horizon BUV395 with an Ex Max of 348-nm and an acceptor dye with an Em Max at 661-nm. BD Horizon Brilliant BUV661 can be excited by the ultraviolet laser (355 nm) and detected with a 670/25 filter and a 630 nm LP. Due to cross laser excitation of this dye, there may be significant spillover into channels detecting APC-like emissions (eg, 670/25-nm filter).
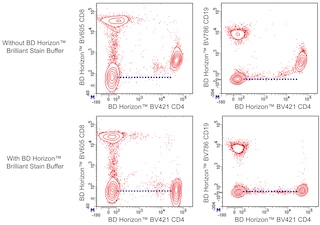
Due to spectral differences between labeled cells and beads, using BD™ CompBeads can result in incorrect spillover values when used with BD Horizon BUV661 reagents. Therefore, the use of BD CompBeads or BD CompBeads Plus to determine spillover values for these reagents is not recommended. Different BUV661 reagents (eg, CD4 vs. CD45) can have slightly different fluorescence spillover therefore, it may also be necessary to use clone-specific compensation controls when using these reagents.