Preparation And Storage
Recommended Assay Procedures
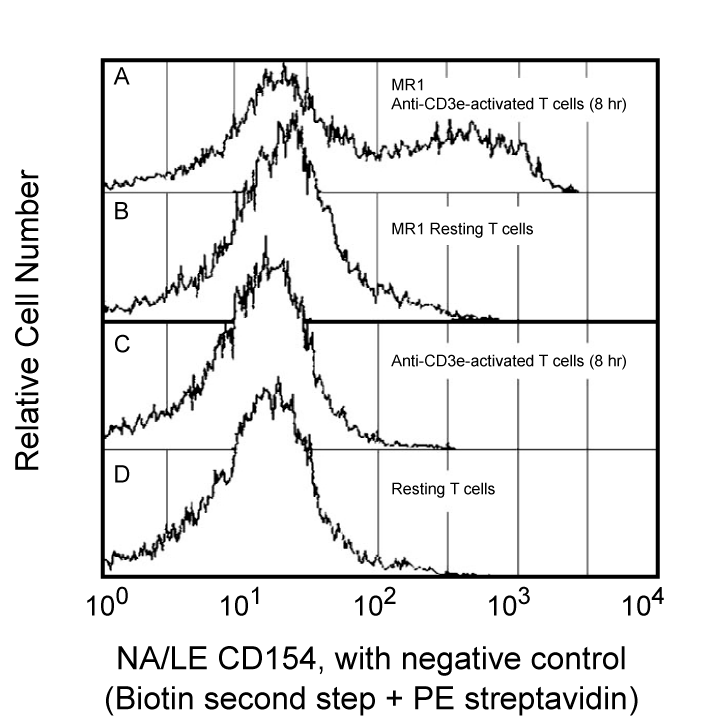
Flow cytometry: For the detection of mouse CD154 on activated peripheral T cells, it is strongly recommended that T cells be purified before activation. Mouse CD154 is transiently expressed on the surfaces of activated normal T cells and certain T cell clones with a maximal level detected 6-8 hours post-activation. Activation with immobilized anti-CD3e mAb (e.g., 145-2C11, Cat. No. 557306/553058, or 500A2, Cat. No. 553238) is sufficient to induce CD154 expression on CD4+ cells. It has been reported that CD8+ cells express CD154 only in response to PMA/Ionomycin treatment. Therefore, for detection of CD154, it is crucial to utilize the proper activation stimuli and to stain cells at the optimal time for CD154 expression. We recommend the use of biotinylated mouse anti-hamster IgG cocktail (Cat. No. 554010) followed by a "bright" second-step reagent, such as Streptavidin-PE (Cat. No. 554061), for optimal detection of CD154.
Product Notices
- Since applications vary, each investigator should titrate the reagent to obtain optimal results.
- Although hamster immunoglobulin isotypes have not been well defined, BD Biosciences Pharmingen has grouped Armenian and Syrian hamster IgG monoclonal antibodies according to their reactivity with a panel of mouse anti-hamster IgG mAbs. A table of the hamster IgG groups, Reactivity of Mouse Anti-Hamster Ig mAbs, may be viewed at http://www.bdbiosciences.com/documents/hamster_chart_11x17.pdf.
- Please refer to www.bdbiosciences.com/us/s/resources for technical protocols.
Companion Products


.png?imwidth=320)
The MR1 monoclonal antibody specifically binds to CD154 (CD40 Ligand, gp39), an accessory molecule expressed on activated T helper (CD4+) lymphocytes. CD154 has also been detected on other types of leukocytes, including CD8+ T cells, medullary thymocytes, activated CD4+ NK-T cells, and human NK cells. CD154 plays an important role in costimulatory interactions between T and B lymphocytes and between antigen-presenting cells and lymphocytes, regulating the immune response at multiple levels. MR1 mAb inhibits in vitro activation of B lymphocytes by T helper cells by blocking interaction of gp39 with CD40. In vitro interactions of T cells and antigen-presenting cells can also be blocked by the MR1 antibody. In vivo treatment with MR1 antibody blocks the development of experimental autoimmune disease, inhibits formation of germinal centers and generation of memory B cells, reduces T-lymphocyte responses to allogeneic cells and allografts, prevents intrathymic deletion of self-reactive T lymphocytes, and disrupts antigen-specific T-cell responses.
Development References (24)
-
Carbone E, Ruggiero G, Terrazzano G, et al. A new mechanism of NK cell cytotoxicity activation: the CD40-CD40 ligand interaction. J Exp Med. 1997; 185(12):2053-2060. (Biology). View Reference
-
DeKruyff RH, Gieni RS, Umetsu DT. Antigen-driven but not lipopolysaccharide-driven IL-12 production in macrophages requires triggering of CD40. J Immunol. 1997; 158(1):359-366. (Clone-specific: Blocking). View Reference
-
Dunn RJ, Luedecker CJ, Haugen HS, Clegg CH, Farr AG. Thymic overexpression of CD40 ligand disrupts normal thymic epithelial organization. J Histochem Cytochem. 1997; 45(1):129-141. (Clone-specific: Immunohistochemistry). View Reference
-
Durie FH, Fava RA, Foy TM, Aruffo A, Ledbetter JA, Noelle RJ. Prevention of collagen-induced arthritis with an antibody to gp39, the ligand for CD40. Science. 1993; 261(5126):1328-1330. (Clone-specific: Blocking). View Reference
-
Foy TM, Laman JD, Ledbetter JA, Aruffo A, Claassen E, Noelle RJ. gp39-CD40 interactions are essential for germinal center formation and the development of B cell memory. J Exp Med. 1994; 180(1):157-163. (Clone-specific: Blocking). View Reference
-
Foy TM, Page DM, Waldschmidt TJ, et al. An essential role for gp39, the ligand for CD40, in thymic selection. J Exp Med. 1995; 182(5):1377-1388. (Clone-specific: Blocking). View Reference
-
Garside P, Ingulli E, Merica RR, Johnson JG, Noelle RJ, Jenkins MK. Visualization of specific B and T lymphocyte interactions in the lymph node. Science. 1998; 281(5373):96-99. (Clone-specific: Blocking). View Reference
-
Graca L, Honey K, Adams E, Cobbold SP, Waldmann H. Cutting edge: anti-CD154 therapeutic antibodies induce infectious transplantation tolerance. J Immunol. 2000; 165(9):4783-4786. (Clone-specific: Blocking). View Reference
-
Grewal IS, Flavell RA. CD40 and CD154 in cell-mediated immunity. Annu Rev Immunol. 1998; 16:111-135. (Biology). View Reference
-
Griggs ND, Agersborg SS, Noelle RJ, Ledbetter JA, Linsley PS, Tung KS. The relative contribution of the CD28 and gp39 costimulatory pathways in the clonal expansion and pathogenic acquisition of self-reactive T cells. J Exp Med. 1996; 183(3):801-810. (Clone-specific: Blocking). View Reference
-
Kalled SL, Cutler AH, Datta SK, Thomas DW. Anti-CD40 ligand antibody treatment of SNF1 mice with established nephritis: preservation of kidney function. J Immunol. 1998; 160(5):2158-2165. (Clone-specific: Blocking). View Reference
-
Kawano T, Cui J, Koezuka Y, et al. CD1d-restricted and TCR-mediated activation of valpha14 NKT cells by glycosylceramides. Science. 1997; 278(5343):1626-1629. (Clone-specific: Blocking). View Reference
-
Kelsall BL, Stuber E, Neurath M, Strober W. Interleukin-12 production by dendritic cells. The role of CD40-CD40L interactions in Th1 T-cell responses. Ann N Y Acad Sci. 1996; 795:116-126. (Clone-specific: Blocking). View Reference
-
Laman JD, Claassen E, Noelle RJ. Functions of CD40 and its ligand, gp39 (CD40L). Crit Rev Immunol. 1996; 16(1):59-108. (Biology). View Reference
-
Larsen CP, Elwood ET, Alexander DZ, et al. Long-term acceptance of skin and cardiac allografts after blocking CD40 and CD28 pathways. Nature. 1996; 381(6581):434-438. (Clone-specific: Blocking). View Reference
-
Lettesjö H, Burd GP, Mageed RA. CD4+ T lymphocytes with constitutive CD40 ligand in preautoimmune (NZB x NZW)F1 lupus-prone mice: phenotype and possible role in autoreactivity. J Immunol. 2000; 165(7):4095-4104. (Clone-specific: Immunohistochemistry). View Reference
-
Masten BJ, Yates JL, Pollard Koga AM, Lipscomb MF. Characterization of accessory molecules in murine lung dendritic cell function: roles for CD80, CD86, CD54, and CD40L. Am J Respir Cell Mol Biol. 1997; 16(3):335-342. (Clone-specific: Blocking). View Reference
-
Miga AJ, Masters SR, Durell BG, et al. Dendritic cell longevity and T cell persistence is controlled by CD154-CD40 interactions. Eur J Immunol. 2001; 31(3):959-965. (Clone-specific: Blocking). View Reference
-
Nishimura T, Kitamura H, Iwakabe K, et al. The interface between innate and acquired immunity: glycolipid antigen presentation by CD1d-expressing dendritic cells to NKT cells induces the differentiation of antigen-specific cytotoxic T lymphocytes. Int Immunol. 2000; 12(7):987-994. (Clone-specific: Blocking). View Reference
-
Noelle RJ, Roy M, Shepherd DM, Stamenkovic I, Ledbetter JA, Aruffo A. A 39-kDa protein on activated helper T cells binds CD40 and transduces the signal for cognate activation of B cells. Proc Natl Acad Sci U S A. 1992; 89(14):6550-6554. (Immunogen: Blocking). View Reference
-
Roy M, Aruffo A, Ledbetter J, Linsley P, Kehry M, Noelle R. Studies on the interdependence of gp39 and B7 expression and function during antigen-specific immune responses. Eur J Immunol. 1995; 25(2):596-603. (Clone-specific: Blocking). View Reference
-
Roy M, Waldschmidt T, Aruffo A, Ledbetter JA, Noelle RJ. The regulation of the expression of gp39, the CD40 ligand, on normal and cloned CD4+ T cells. J Immunol. 1993; 151(5):2497-2510. (Biology). View Reference
-
Tian L, Noelle RJ, Lawrence DA. Activated T cells enhance nitric oxide production by murine splenic macrophages through gp39 and LFA-1. Eur J Immunol. 1995; 25(1):306-309. (Clone-specific: Blocking). View Reference
-
Tomura M, Yu WG, Ahn HJ, et al. A novel function of Valpha14+CD4+NKT cells: stimulation of IL-12 production by antigen-presenting cells in the innate immune system. J Immunol. 1999; 163(1):93-101. (Biology). View Reference
Please refer to Support Documents for Quality Certificates
Global - Refer to manufacturer's instructions for use and related User Manuals and Technical data sheets before using this products as described
Comparisons, where applicable, are made against older BD Technology, manual methods or are general performance claims. Comparisons are not made against non-BD technologies, unless otherwise noted.
For Research Use Only. Not for use in diagnostic or therapeutic procedures.